

Alien, a highly conserved protein with characteristics of a corepressor for members of the nuclear hormone receptor superfamily
- PMID: 10207062
- PMCID: PMC84131
- DOI: 10.1128/MCB.19.5.3383
Alien, a highly conserved protein with characteristics of a corepressor for members of the nuclear hormone receptor superfamily
Abstract
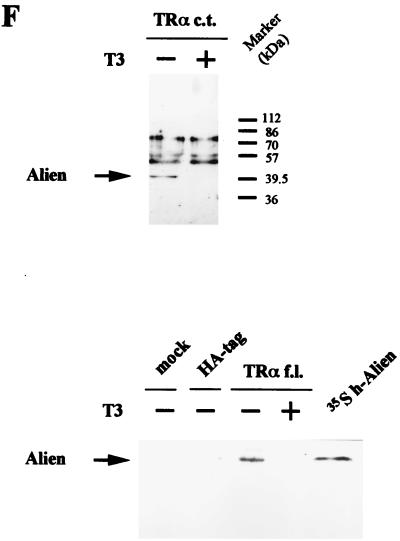
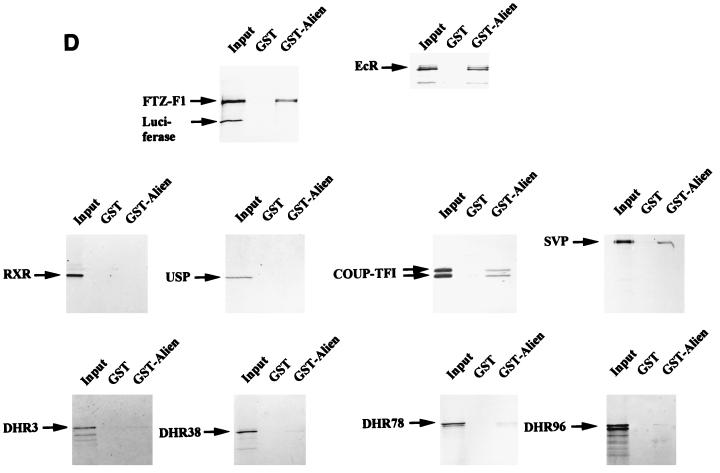
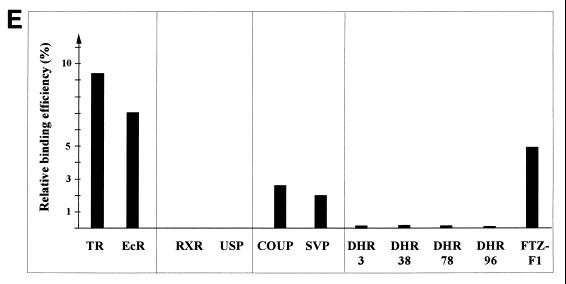
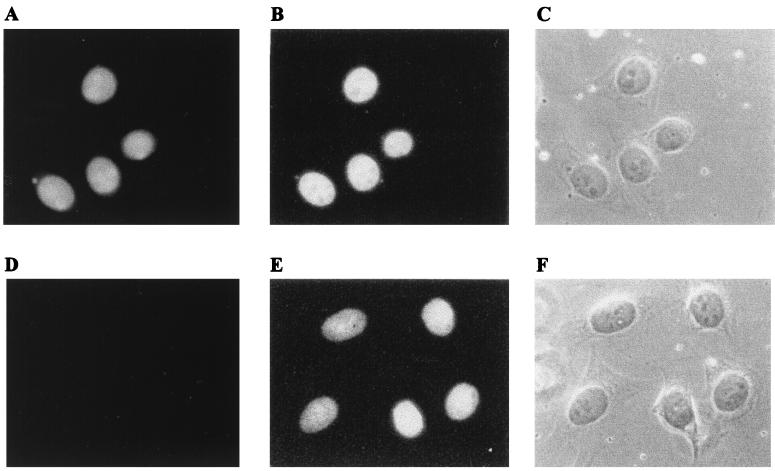
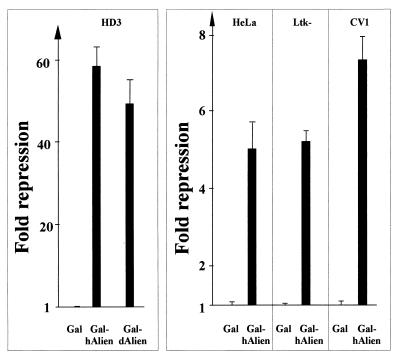
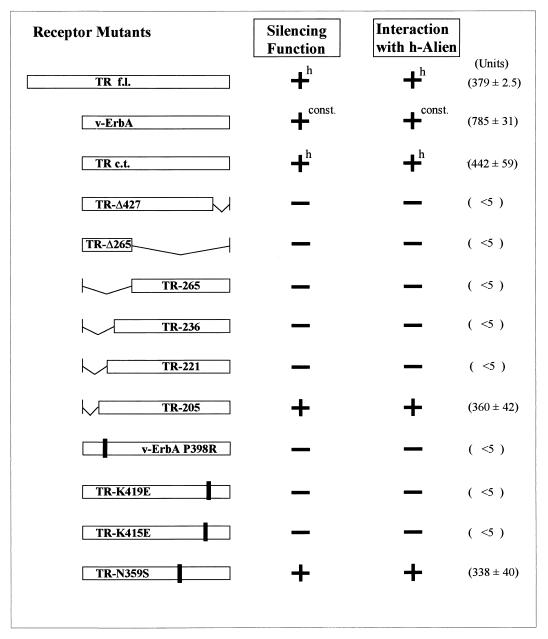
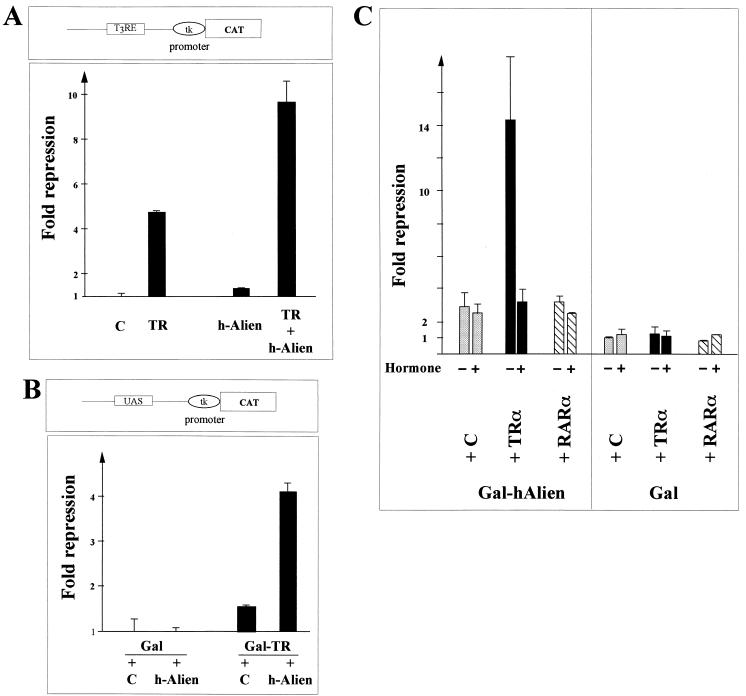
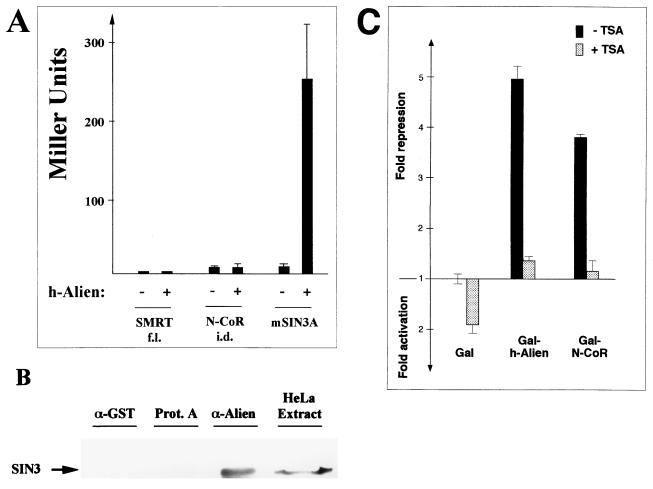
Some members of nuclear hormone receptors, such as the thyroid hormone receptor (TR), silence gene expression in the absence of the hormone. Corepressors, which bind to the receptor's silencing domain, are involved in this repression. Hormone binding leads to dissociation of corepressors and binding of coactivators, which in turn mediate gene activation. Here, we describe the characteristics of Alien, a novel corepressor. Alien interacts with TR only in the absence of hormone. Addition of thyroid hormone leads to dissociation of Alien from the receptor, as shown by the yeast two-hybrid system, glutathione S-transferase pull-down, and coimmunoprecipitation experiments. Reporter assays indicate that Alien increases receptor-mediated silencing and that it harbors an autonomous silencing function. Immune staining shows that Alien is localized in the cell nucleus. Alien is a highly conserved protein showing 90% identity between human and Drosophila. Drosophila Alien shows similar activities in that it interacts in a hormone-sensitive manner with TR and harbors an autonomous silencing function. Specific interaction of Alien is seen with Drosophila nuclear hormone receptors, such as the ecdysone receptor and Seven-up, the Drosophila homologue of COUP-TF1, but not with retinoic acid receptor, RXR/USP, DHR 3, DHR 38, DHR 78, or DHR 96. These properties, taken together, show that Alien has the characteristics of a corepressor. Thus, Alien represents a member of a novel class of corepressors specific for selected members of the nuclear hormone receptor superfamily.
Figures

References
-
- Alland L, Muhle R, Hou H, Jr, Potes J, Chin L, Schreiber-Agus N, DePinho R A. Role of N-CoR and histone deacetylase in SIN3-mediated transcriptional repression. Nature. 1997;387:49–55. - PubMed
-
- Ayer D E, Lawrence Q A, Eisenman R N. Mad-Max transcriptional repression is mediated by ternary complex formation with mammalian homologs of yeast repressor Sin3. Cell. 1995;80:767–776. - PubMed
-
- Baniahmad A, Steiner C, Köhne A C, Renkawitz R. Modular structure of a chicken lysozyme silencer: involvement of an unusual thyroid hormone receptor binding site. Cell. 1990;61:505–514. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases