

The Saccharomyces cerevisiae homologues of endonuclease III from Escherichia coli, Ntg1 and Ntg2, are both required for efficient repair of spontaneous and induced oxidative DNA damage in yeast
- PMID: 10207101
- PMCID: PMC84209
- DOI: 10.1128/MCB.19.5.3779
The Saccharomyces cerevisiae homologues of endonuclease III from Escherichia coli, Ntg1 and Ntg2, are both required for efficient repair of spontaneous and induced oxidative DNA damage in yeast
Abstract
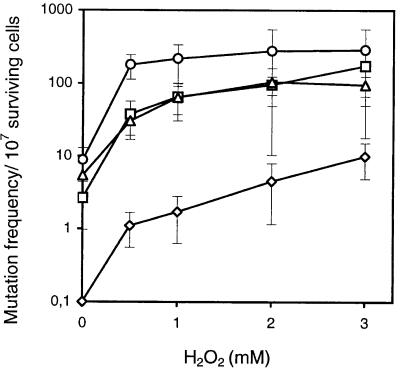
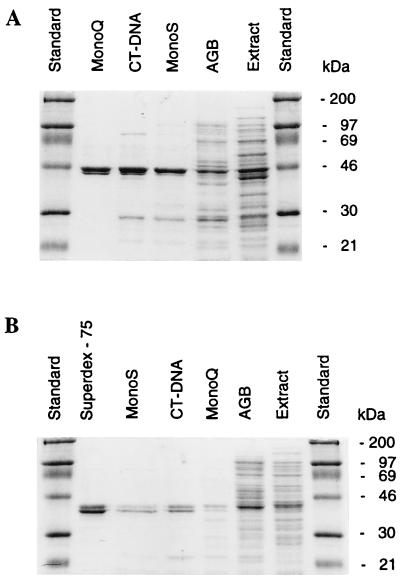
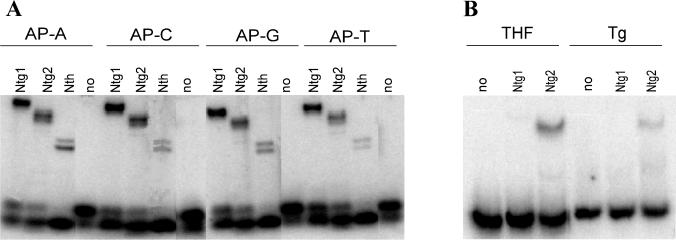
Endonuclease III from Escherichia coli is the prototype of a ubiquitous DNA repair enzyme essential for the removal of oxidized pyrimidine base damage. The yeast genome project has revealed the presence of two genes in Saccharomyces cerevisiae, NTG1 and NTG2, encoding proteins with similarity to endonuclease III. Both contain the highly conserved helix-hairpin-helix motif, whereas only one (Ntg2) harbors the characteristic iron-sulfur cluster of the endonuclease III family. We have characterized these gene functions by mutant and enzyme analysis as well as by gene expression and intracellular localization studies. Targeted gene disruption of NTG1 and NTG2 produced mutants with greatly increased spontaneous and hydrogen peroxide-induced mutation frequency relative to the wild type, and the mutation response was further increased in the double mutant. Both enzymes were found to remove thymine glycol and 2, 6-diamino-4-hydroxy-5-N-methylformamidopyrimidine (faPy) residues from DNA with high efficiency. However, on UV-irradiated DNA, saturating concentrations of Ntg2 removed only half of the cytosine photoproducts released by Ntg1. Conversely, 5-hydroxycytosine was removed efficiently only by Ntg2. The enzymes appear to have different reaction modes, as judged from much higher affinity of Ntg2 for damaged DNA and more efficient borhydride trapping of Ntg1 to abasic sites in DNA despite limited DNA binding. Northern blot and promoter fusion analysis showed that NTG1 is inducible by cell exposure to DNA-damaging agents, whereas NTG2 is constitutively expressed. Ntg2 appears to be a nuclear enzyme, whereas Ntg1 was sorted both to the nucleus and to the mitochondria. We conclude that functions of both NTG1 and NTG2 are important for removal of oxidative DNA damage in yeast.
Figures






References
-
- Augeri L, Lee L, Barton A B, Doetsch P W. Purification, characterization, gene cloning, and expression of Saccharomyces cerevisiae redoxyendonuclease, a homolog of Escherichia coli endonuclease III. Biochemistry. 1997;36:721–729. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials