

A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development
- PMID: 10215629
- PMCID: PMC316641
- DOI: 10.1101/gad.13.8.1025
A Cbfa1-dependent genetic pathway controls bone formation beyond embryonic development
Abstract
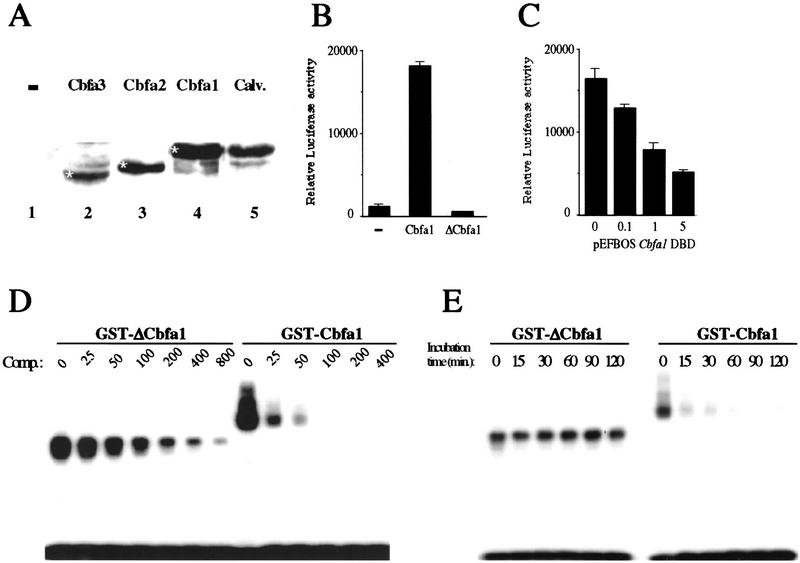
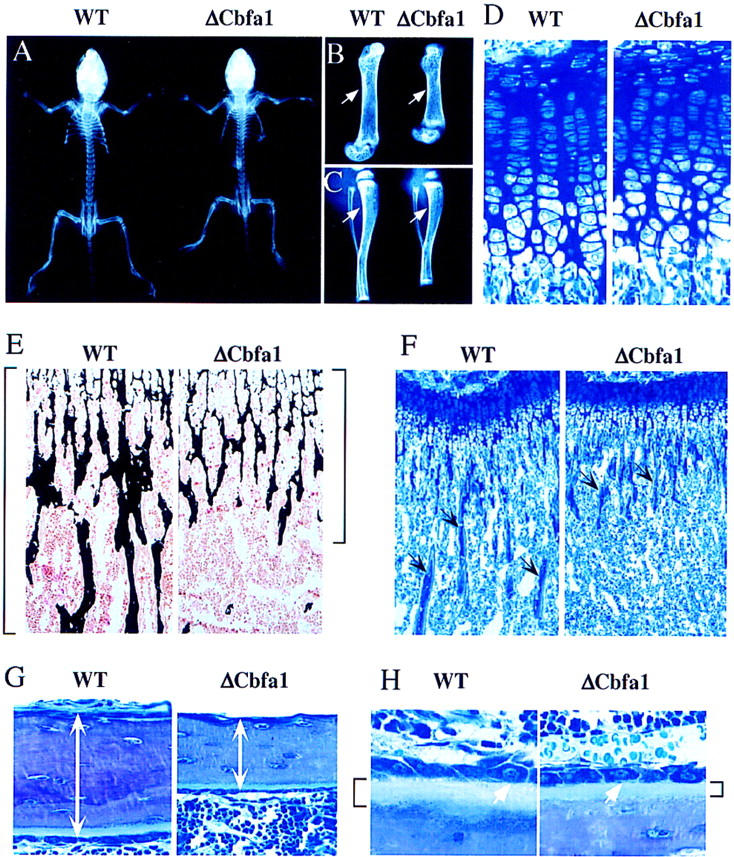
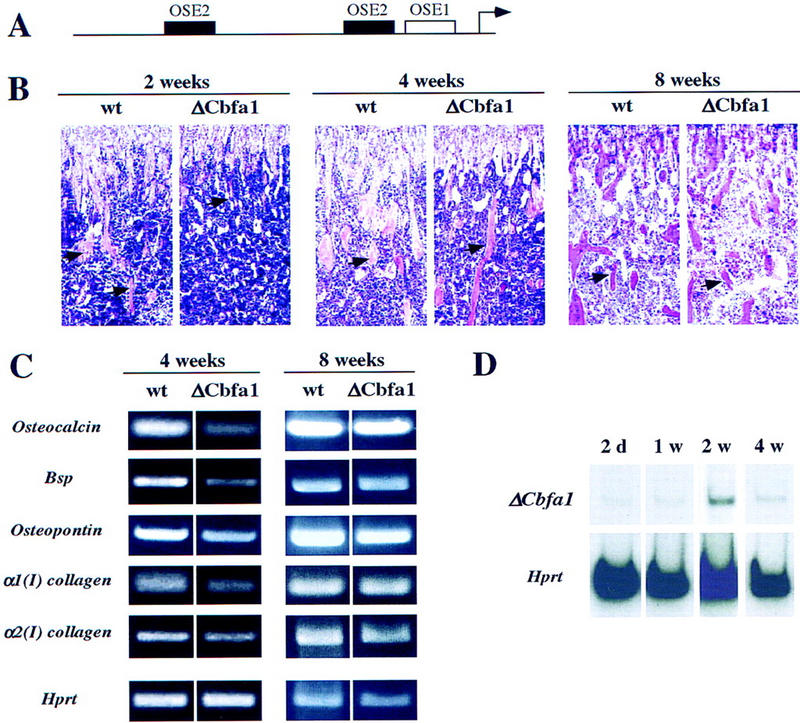
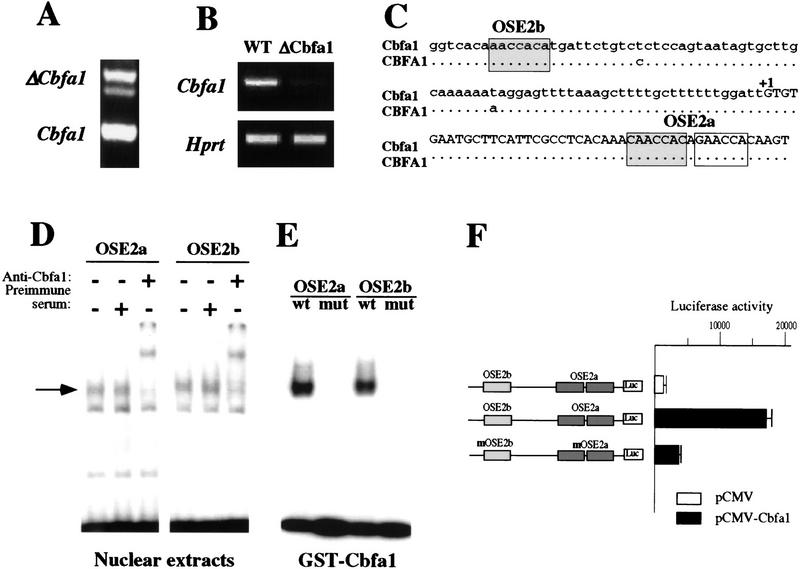
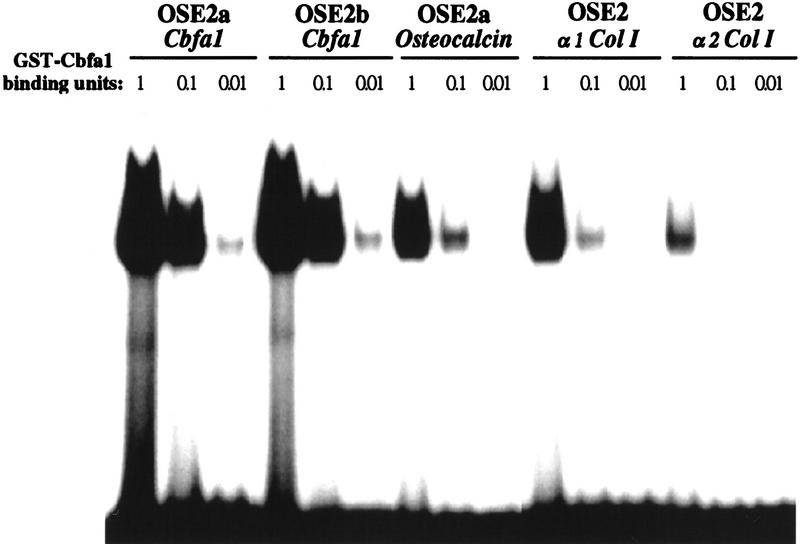
The molecular mechanisms controlling bone extracellular matrix (ECM) deposition by differentiated osteoblasts in postnatal life, called hereafter bone formation, are unknown. This contrasts with the growing knowledge about the genetic control of osteoblast differentiation during embryonic development. Cbfa1, a transcriptional activator of osteoblast differentiation during embryonic development, is also expressed in differentiated osteoblasts postnatally. The perinatal lethality occurring in Cbfa1-deficient mice has prevented so far the study of its function after birth. To determine if Cbfa1 plays a role during bone formation we generated transgenic mice overexpressing Cbfa1 DNA-binding domain (DeltaCbfa1) in differentiated osteoblasts only postnatally. DeltaCbfa1 has a higher affinity for DNA than Cbfa1 itself, has no transcriptional activity on its own, and can act in a dominant-negative manner in DNA cotransfection assays. DeltaCbfa1-expressing mice have a normal skeleton at birth but develop an osteopenic phenotype thereafter. Dynamic histomorphometric studies show that this phenotype is caused by a major decrease in the bone formation rate in the face of a normal number of osteoblasts thus indicating that once osteoblasts are differentiated Cbfa1 regulates their function. Molecular analyses reveal that the expression of the genes expressed in osteoblasts and encoding bone ECM proteins is nearly abolished in transgenic mice, and ex vivo assays demonstrated that DeltaCbfa1-expressing osteoblasts were less active than wild-type osteoblasts. We also show that Cbfa1 regulates positively the activity of its own promoter, which has the highest affinity Cbfa1-binding sites characterized. This study demonstrates that beyond its differentiation function Cbfa1 is the first transcriptional activator of bone formation identified to date and illustrates that developmentally important genes control physiological processes postnatally.
Figures





References
-
- Aubin JE, Liu F. The osteoblast lineage. In: Bilezikian JP, Raisz LG, Rodan GA, editors. Principles of bone biology. San Diego, CA: Academic Press; 1996. pp. 51–67.
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. New York, NY: John Wiley & Sons; 1995.
-
- Bancroft JD, Stevens A. Theory and practice of histological techniques. New York, NY: Churchil Livingston; 1996.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases