

The C-terminal sequence of the lambda holin constitutes a cytoplasmic regulatory domain
- PMID: 10217787
- PMCID: PMC93738
- DOI: 10.1128/JB.181.9.2922-2929.1999
The C-terminal sequence of the lambda holin constitutes a cytoplasmic regulatory domain
Abstract
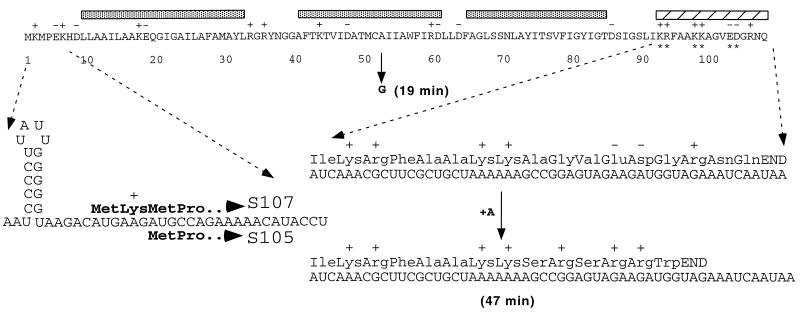
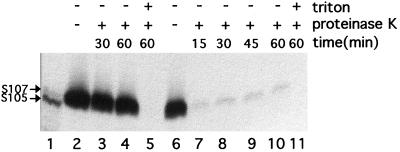
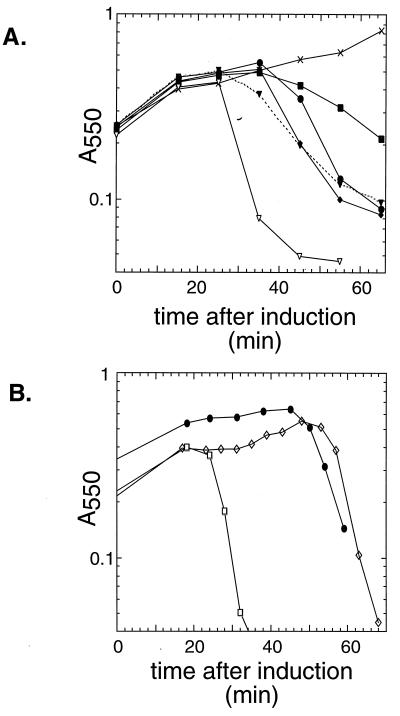
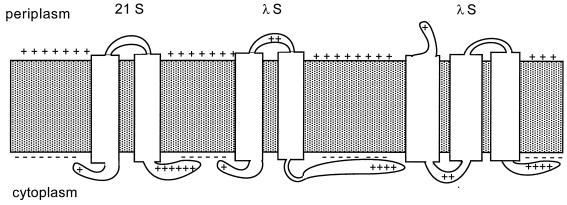
The C-terminal domains of holins are highly hydrophilic and contain clusters of consecutive basic and acidic residues, with the overall net charge predicted to be positive. The C-terminal domain of lambda S was found to be cytoplasmic, as defined by protease accessibility in spheroplasts and inverted membrane vesicles. C-terminal nonsense mutations were constructed in S and found to be lysis proficient, as long as at least one basic residue is retained at the C terminus. In general, the normal intrinsic scheduling of S function is deranged, resulting in early lysis. However, the capacity of each truncated lytic allele for inhibition by the S107 inhibitor product of S is retained. The K97am allele, when incorporated into the phage context, confers a plaque-forming defect because its early lysis significantly reduces the burst size. Finally, a C-terminal frameshift mutation was isolated as a suppressor of the even more severe early lysis defect of the mutant SA52G, which causes lysis at or before the time when the first phage particle is assembled in the cell. This mutation scrambles the C-terminal sequence of S, resulting in a predicted net charge increase of +4, and retards lysis by about 30 min, thus permitting a viable quantity of progeny to accumulate. Thus, the C-terminal domain is not involved in the formation of the lethal membrane lesion nor in the "dual-start" regulation conserved in lambdoid holins. Instead, the C-terminal sequence defines a cytoplasmic regulatory domain which affects the timing of lysis. Comparison of the C-terminal sequences of within holin families suggests that these domains have little or no structure but act as reservoirs of charged residues that interact with the membrane to effect proper lysis timing.
Figures


References
-
- Barenboim, M., C.-Y. Chang, F. dib Hajj, and R. Young. A holin and its inhibitor are both encoded by the phage 21 S gene. Mol. Microbiol., in press.
-
- Bienkowska-Szewczyk K, Lipinska B, Taylor A. The R gene product of bacteriophage λ is the murein transglycosylase. Mol Gen Genet. 1981;184:111–114. - PubMed
-
- Bläsi, U., C.-Y. Chang, and R. Young. Unpublished data.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
