

T cell factor-activated transcription is not sufficient to induce anchorage-independent growth of epithelial cells expressing mutant beta-catenin
- PMID: 10220399
- PMCID: PMC21797
- DOI: 10.1073/pnas.96.9.4947
T cell factor-activated transcription is not sufficient to induce anchorage-independent growth of epithelial cells expressing mutant beta-catenin
Abstract
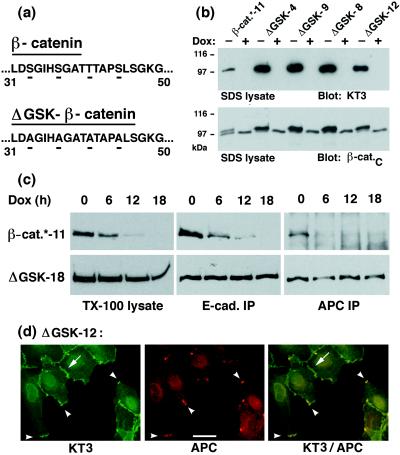
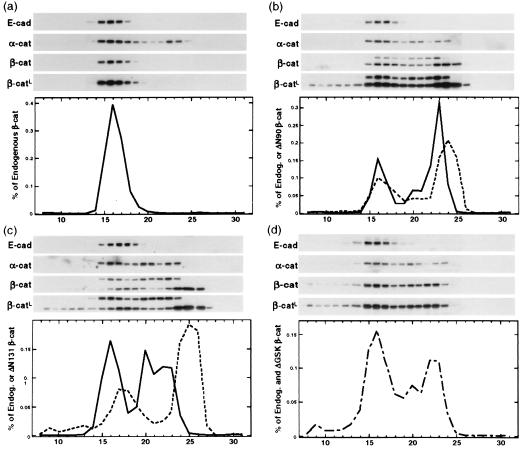
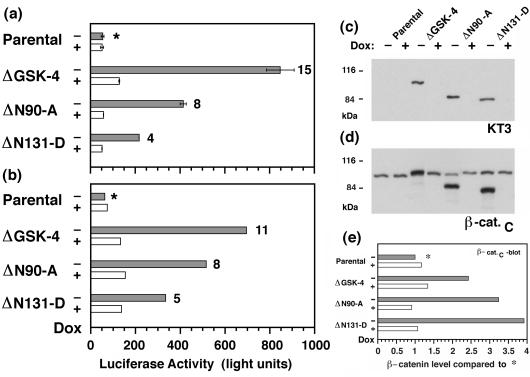
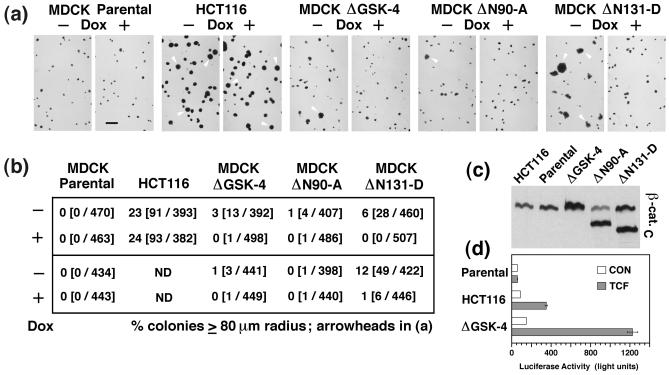
N-terminal mutations in beta-catenin that inhibit beta-catenin degradation are found in primary tumors and cancer cell lines, and increased beta-catenin/T cell factor (TCF)-activated transcription in these cells has been correlated with cancer formation. However, the role of mutant beta-catenin in cell transformation is poorly understood. Here, we compare the ability of different N-terminal mutations of beta-catenin (DeltaN131, DeltaN90, DeltaGSK) to induce TCF-activated transcription and anchorage-independent growth in Madin-Darby canine kidney epithelial cells. Expression of DeltaN90 or DeltaGSK beta-catenin increased TCF-activated transcription but did not induce significant anchorage-independent cell growth. In contrast, deletion of the alpha-catenin-binding site in DeltaN131 beta-catenin reduced TCF-activated transcription, compared with that induced by DeltaN90 or DeltaGSK beta-catenin, but significantly enhanced anchorage-independent cell growth.
Figures
References
-
- Gumbiner B M. Curr Opin Cell Biol. 1995;7:634–640. - PubMed
-
- Peifer M. Trends Cell Biol. 1995;5:224–229. - PubMed
-
- Barth A I, Näthke I S, Nelson W J. Curr Opin Cell Biol. 1997;9:683–690. - PubMed
-
- Aberle, H., Butz, S., Stappert, J., Weissig, H., Kemler, R. & Hoschuetzky, H. (1994) J. Cell Sci. 3655–3663. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
