

Identification of profilin and src homology 3 domains as binding partners for Drosophila enabled
- PMID: 10220404
- PMCID: PMC21802
- DOI: 10.1073/pnas.96.9.4977
Identification of profilin and src homology 3 domains as binding partners for Drosophila enabled
Abstract
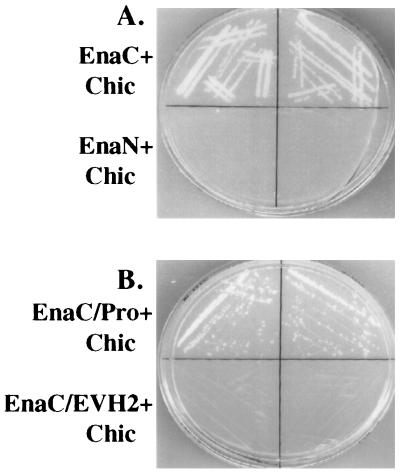
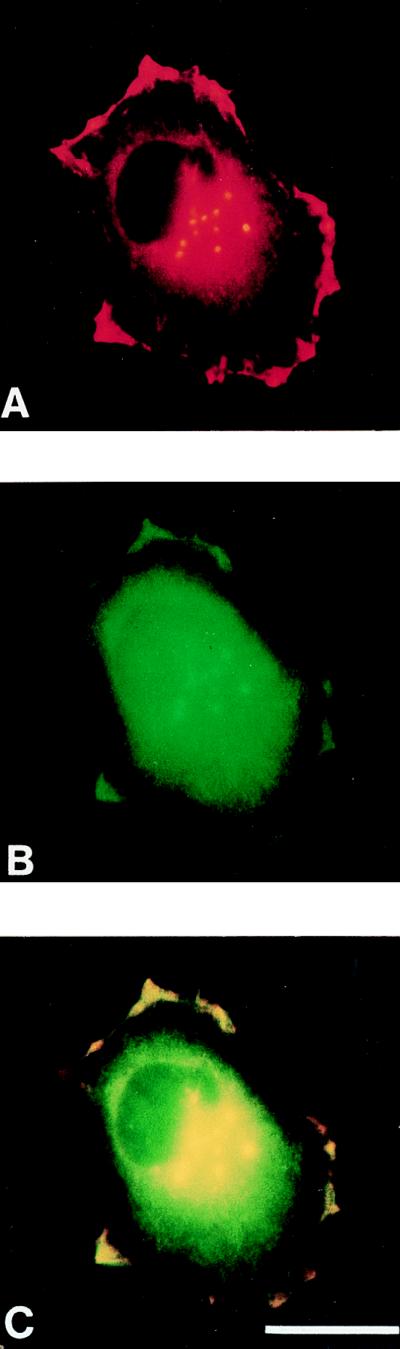
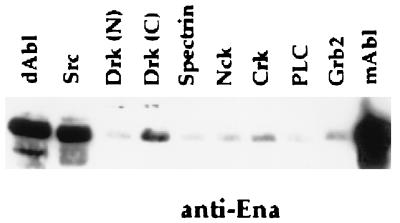
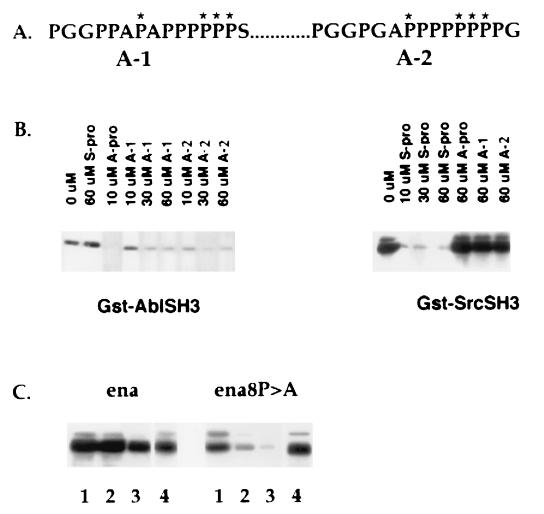
Drosophila Enabled (Ena) was first identified as a genetic suppressor of mutations in the Abelson tyrosine kinase and subsequently was shown to be a member of the Ena/vasodilator-stimulated phosphoprotein family of proteins. All members of this family have a conserved domain organization, bind the focal adhesion protein zyxin, and localize to focal adhesions and stress fibers. Members of this family are thought to be involved in the regulation of cytoskeleton dynamics. The Ena protein sequence has multiple poly-(L-proline) residues with similarity to both profilin and src homology 3 binding sites. Here, we show that Ena can bind directly to the Drosophila homolog of profilin, chickadee. Furthermore, Ena and profilin were colocalized in spreading cultured cells. We report that the proline-rich region of Ena is responsible for this interaction as well as for mediating binding to the src homology 3 domain of the Abelson tyrosine kinase. These data support the hypothesis that Ena provides a regulated link between signal transduction and cytoskeleton assembly in the developing Drosophila embryo.
Figures


References
-
- Carlier M F. Curr Opin Cell Biol. 1998;10:45–51. - PubMed
-
- Dramsi S, Cossart P. Annu Rev Cell Dev Biol. 1998;14:137–166. - PubMed
-
- Elch M D, Mallavarapu A, Rosenblat J, Mitchison T J. Curr Opin Cell Biol. 1997;9:54–61. - PubMed
-
- Gertler F, Niebuhr K, Reinhard M, Wehland J, Soriano P. Cell. 1996;87:227–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
