

Replication of Toxoplasma gondii, but not Trypanosoma cruzi, is regulated in human fibroblasts activated with gamma interferon: requirement of a functional JAK/STAT pathway
- PMID: 10225879
- PMCID: PMC115962
- DOI: 10.1128/IAI.67.5.2233-2240.1999
Replication of Toxoplasma gondii, but not Trypanosoma cruzi, is regulated in human fibroblasts activated with gamma interferon: requirement of a functional JAK/STAT pathway
Abstract
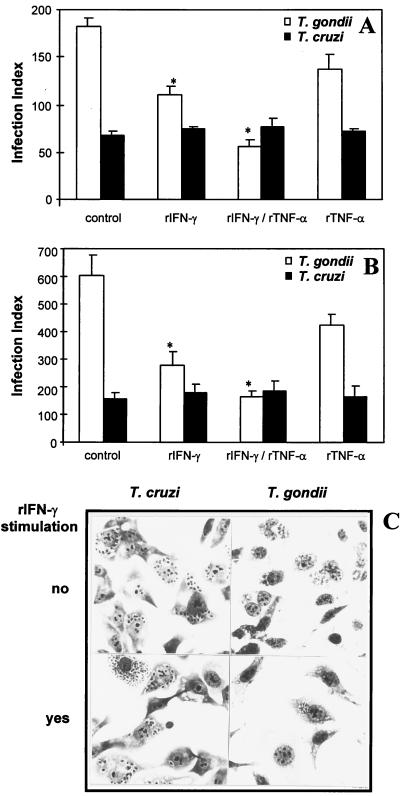
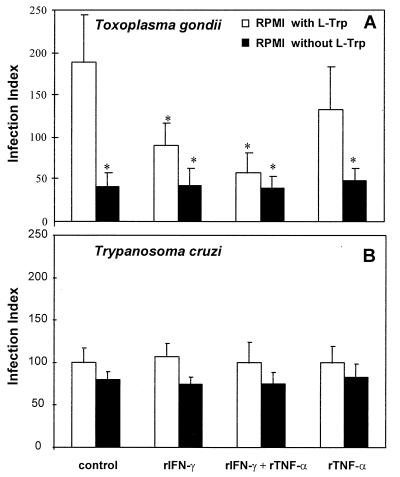
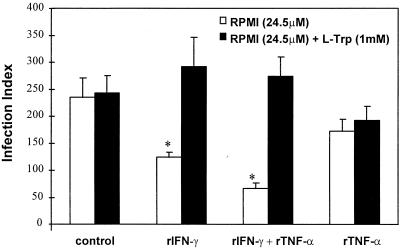
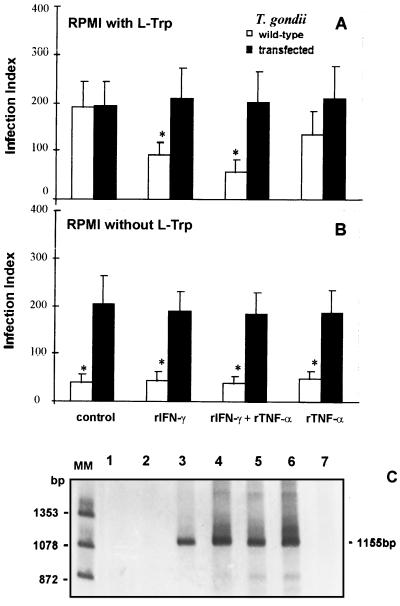
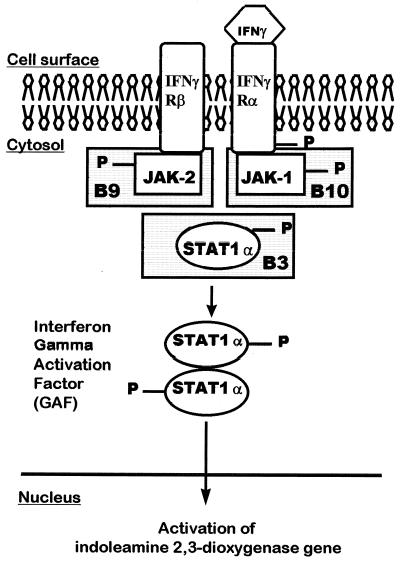
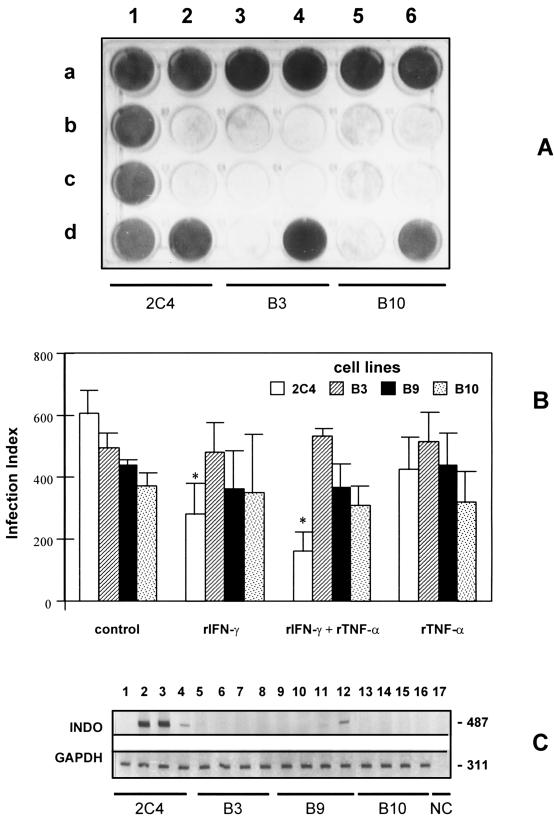
To study the role of tryptophan degradation by indoleamine 2, 3-dioxygenase (INDO) in the control of Trypanosoma cruzi or Toxoplasma gondii replication, we used human fibroblasts and a fibrosarcoma cell line (2C4). The cells were cultured in the presence or absence of recombinant gamma interferon (rIFN-gamma) and/or recombinant tumor necrosis factor alpha (rTNF-alpha) for 24 h and were then infected with either T. cruzi or T. gondii. Intracellular parasite replication was evaluated 24 or 48 h after infection. Treatment with rIFN-gamma and/or rTNF-alpha had no inhibitory effect on T. cruzi replication. In contrast, 54, 73, or 30% inhibition of T. gondii replication was observed in the cells treated with rIFN-gamma alone, rIFN-gamma plus rTNF-alpha, or TNF-alpha alone, respectively. The replication of T. gondii tachyzoites in cytokine-activated cells was restored by the addition of extra tryptophan to the culture medium. Similarly, T. gondii tachyzoites transfected with bacterial tryptophan synthase were not sensitive to the microbiostatic effect of rIFN-gamma. We also investigated the basis of the cytokine effect on parasite replication by using the three mutant cell lines B3, B9, and B10 derived from 2C4 and expressing defective STAT1alpha (signal transducer and activator of transcription), JAK2 (Janus family of cytoplasmic tyrosine kinases), or JAK1, respectively, three important elements of a signaling pathway triggered by rIFN-gamma. We found that rTNF-alpha was able to induce low levels expression of INDO mRNA in the parental cell line, as well as the cell line lacking functional JAK2. In contrast to the parental cell line (2C4), rIFN-gamma was not able to induce the expression of INDO mRNA or microbiostatic activity in any of the mutant cell lines. These findings indicate the essential requirement of the JAK/STAT pathway for the induction of high levels of INDO mRNA, tryptophan degradation, and the anti-Toxoplasma activity inside human nonprofessional phagocytic cells.
Figures
References
-
- Adams L B, Hibbs J B, Taintor R R, Krahenbuhl J L. Microbiostatic effect of murine activated macrophages for Toxoplasma gondii: role for synthesis of inorganic nitrogen oxides from l-arginine. J Immunol. 1990;144:2725–2729. - PubMed
-
- Andrews N W. Living dangerously: how Trypanosoma cruzi uses lysosomes to get inside host cells, and then escapes into the cytoplasm. Biol Res. 1993;26:65–67. - PubMed
-
- Bertelli M S M, Golgher R R, Brener Z. Intraspecific variation in Trypanosoma cruzi: effect of temperature on the intracellular differentiation in tissue culture. J Parasitol. 1977;63:434–437. - PubMed
-
- Bonjardim C A. JAK/STAT deficient cell lines. Braz J Med Biol Res. 1998;31:1389–1395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
