

Molecular interactions of Porphyromonas gingivalis fimbriae with host proteins: kinetic analyses based on surface plasmon resonance
- PMID: 10225901
- PMCID: PMC115984
- DOI: 10.1128/IAI.67.5.2399-2405.1999
Molecular interactions of Porphyromonas gingivalis fimbriae with host proteins: kinetic analyses based on surface plasmon resonance
Abstract
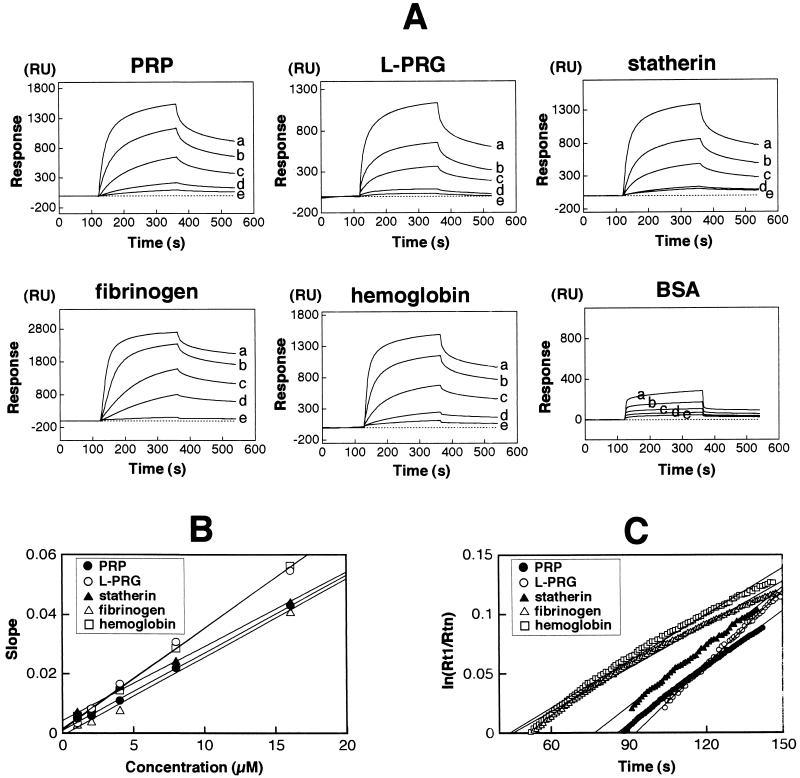
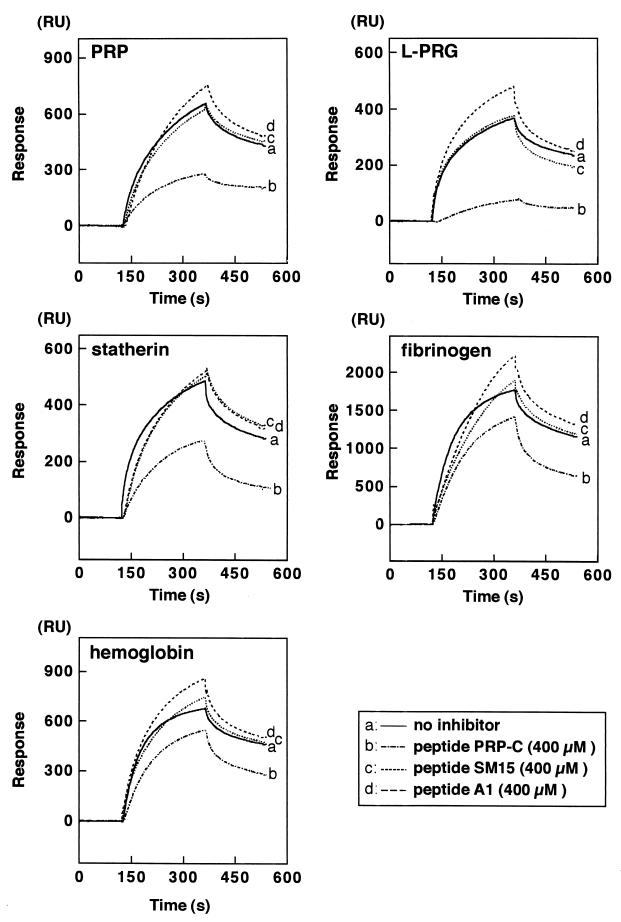
Fimbriae of Porphyromonas gingivalis are thought to play an important role in the colonization and invasion of periodontal tissues. In this study, we analyzed the interactions of P. gingivalis fimbriae with human hemoglobin, fibrinogen, and salivary components (i.e., proline-rich protein [PRP], proline-rich glycoprotein [PRG], and statherin) based on surface plasmon resonance (SPR) spectroscopy with a biomolecular interaction analyzing system (BIAcore). The real-time observation showed that the fimbriae interacted more quickly with hemoglobin and PRG than with other proteins and more intensely with fibrinogen. The significant association constant (ka) values obtained by BIAcore demonstrated that the interactions between fimbriae and these host proteins are specific. These estimated Ka values were not too different; however, the Ka values for hemoglobin (2.43 x 10(6)) and fibrinogen (2.16 x 10(6)) were statistically greater than those for the salivary proteins (1.48 x 10(6) to 1.63 x 10(6)). The Ka value of anti-fimbriae immunoglobulin G for fimbriae was estimated to be 1. 22 x 10(7), which was 6.55-fold higher than the mean Ka value of the host proteins. Peptide PRP-C, a potent inhibitor of PRP-fimbriae interaction, dramatically inhibited fimbrial association to PRP and PRG and was also inhibitory against other host proteins by BIAcore. The binding of fimbriae to these proteins was also evaluated by other methods with hydroxyapatite beads or polystyrene microtiter plates. The estimated binding abilities differed considerably, depending on the assay method that was used. It was noted that the binding capacity of PRP was strongly diminished by immobilization on a polystyrene surface. Taken together, these findings suggest that P. gingivalis fimbriae possess a strong ability to interact with the host proteins which promote bacterial adherence to the oral cavity and that SPR spectroscopy is a useful method for analyzing specific protein-fimbriae interactions.
Figures
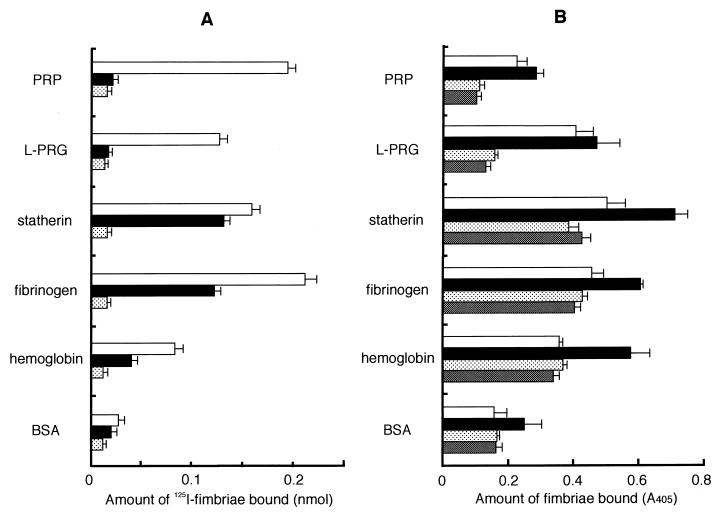
 ). The specific
binding level was calculated by subtracting the nonspecific binding
level. Significant differences (P < 0.01) were
observed among fimbrial binding levels (□) to host proteins in the
absence of peptide PRP-C. (B) Binding assay performed with polystyrene
surface. The wells of 96-well microtiter plates were coated with host
proteins or BSA (100 μl of 0.1 mg/ml in PBS) at 37°C for 2 h.
An aliquot of fimbriae and, if necessary, peptide PRP-C as an inhibitor
was added to the wells and then incubated at RT for 1 h. Rabbit
anti-fimbriae immunoglobulin G (1:1,000) was added to detect the amount
of bound fimbriae. □, 12.2 pmol of added fimbriae; ■, 24.4 pmol of
added fimbriae; , 12.2 pmol of fimbriae plus 100 pmol of added
peptide PRP-C; ▩, 12.2 pmol of fimbriae plus 1 nmol of added peptide
PRP-C. Significant differences (P < 0.01) were
observed among fimbrial binding levels (■) to host proteins in the
absence of peptide PRP-C and between PRP and BSA and between fibrinogen
and hemoglobin. All assays were performed in triplicate, on three
separate occasions.
). The specific
binding level was calculated by subtracting the nonspecific binding
level. Significant differences (P < 0.01) were
observed among fimbrial binding levels (□) to host proteins in the
absence of peptide PRP-C. (B) Binding assay performed with polystyrene
surface. The wells of 96-well microtiter plates were coated with host
proteins or BSA (100 μl of 0.1 mg/ml in PBS) at 37°C for 2 h.
An aliquot of fimbriae and, if necessary, peptide PRP-C as an inhibitor
was added to the wells and then incubated at RT for 1 h. Rabbit
anti-fimbriae immunoglobulin G (1:1,000) was added to detect the amount
of bound fimbriae. □, 12.2 pmol of added fimbriae; ■, 24.4 pmol of
added fimbriae; , 12.2 pmol of fimbriae plus 100 pmol of added
peptide PRP-C; ▩, 12.2 pmol of fimbriae plus 1 nmol of added peptide
PRP-C. Significant differences (P < 0.01) were
observed among fimbrial binding levels (■) to host proteins in the
absence of peptide PRP-C and between PRP and BSA and between fibrinogen
and hemoglobin. All assays were performed in triplicate, on three
separate occasions.References
-
- Abraham R, Buxbaum S, Link J, Smith R, Venti C, Darsley M. Screening and kinetic analysis of recombinant anti-CEA antibody fragments. J Immunol Methods. 1995;183:119–125. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
