

Light-dependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis
- PMID: 10225946
- PMCID: PMC2185089
- DOI: 10.1083/jcb.145.3.437
Light-dependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis
Abstract
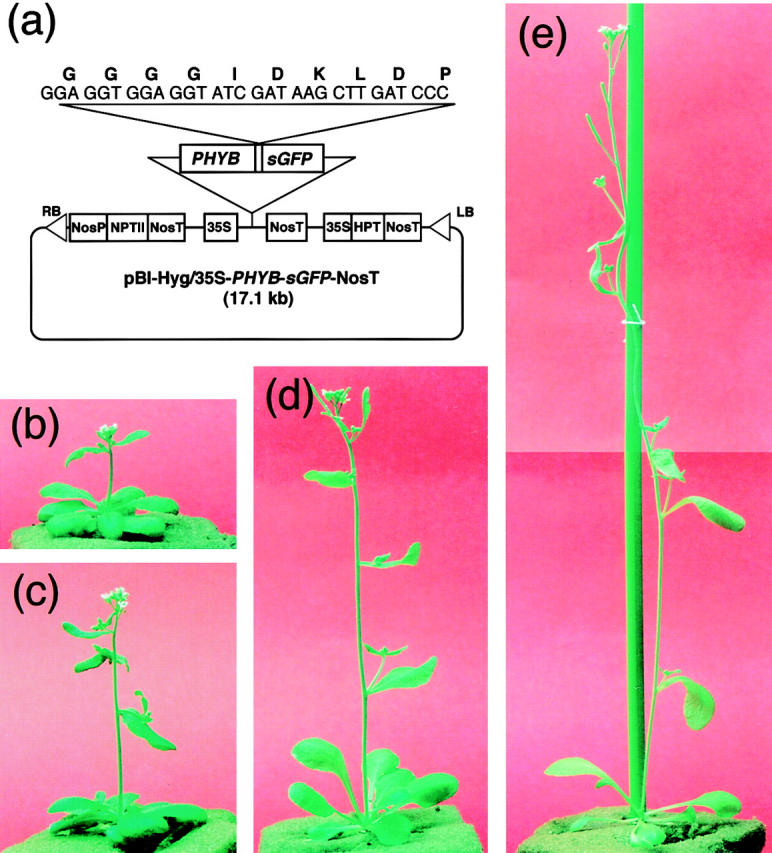
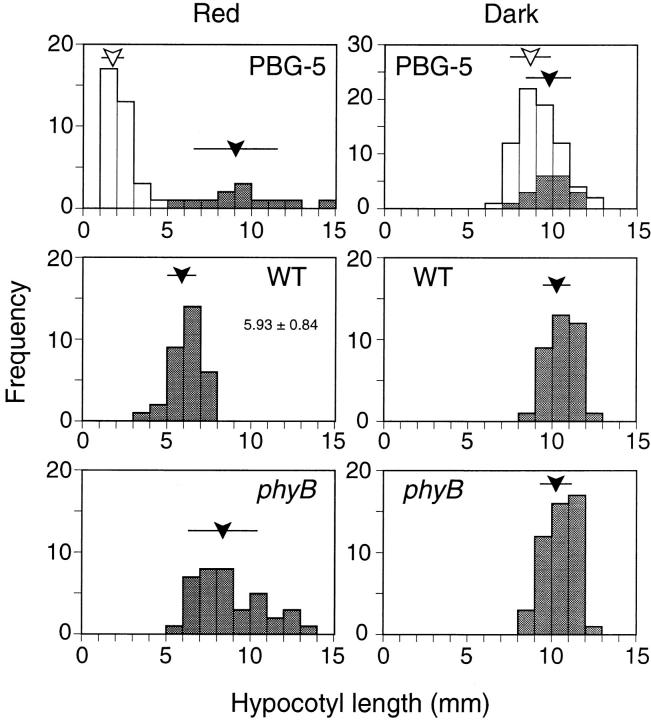
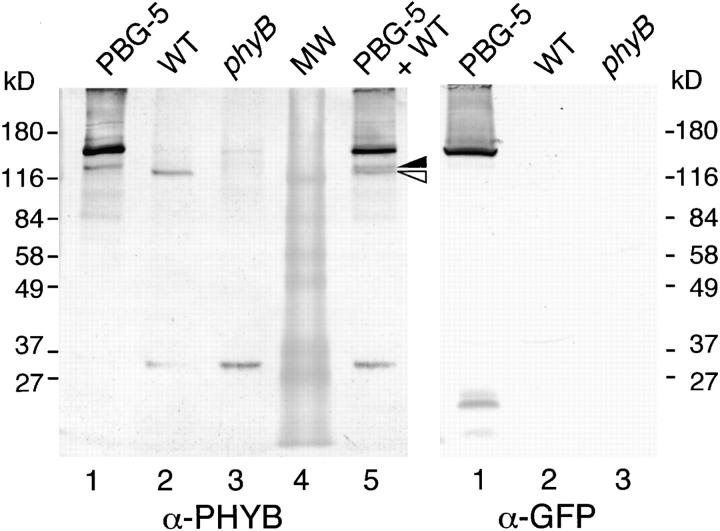
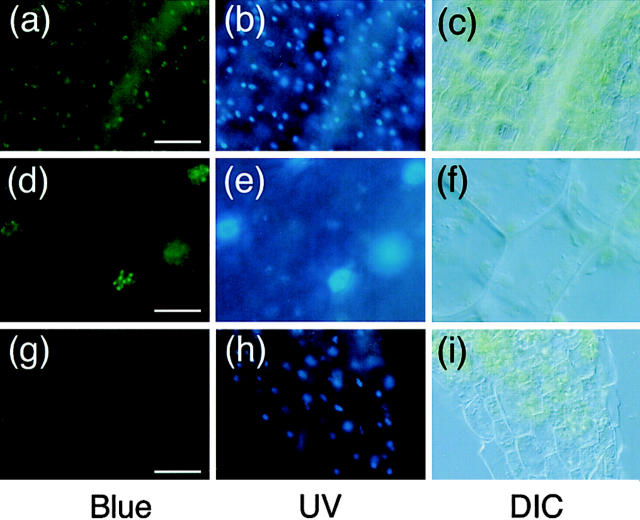
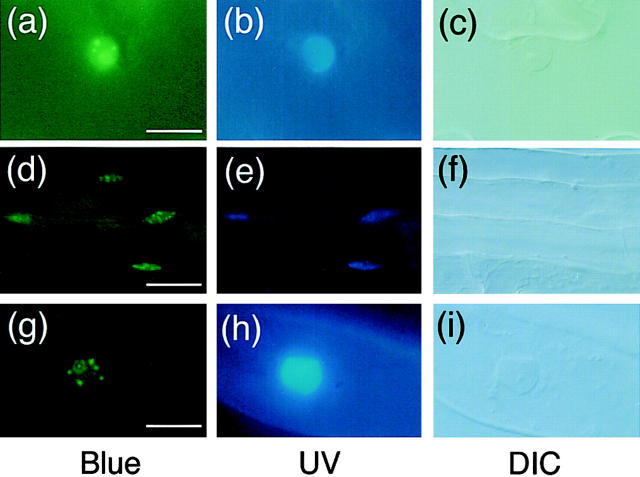
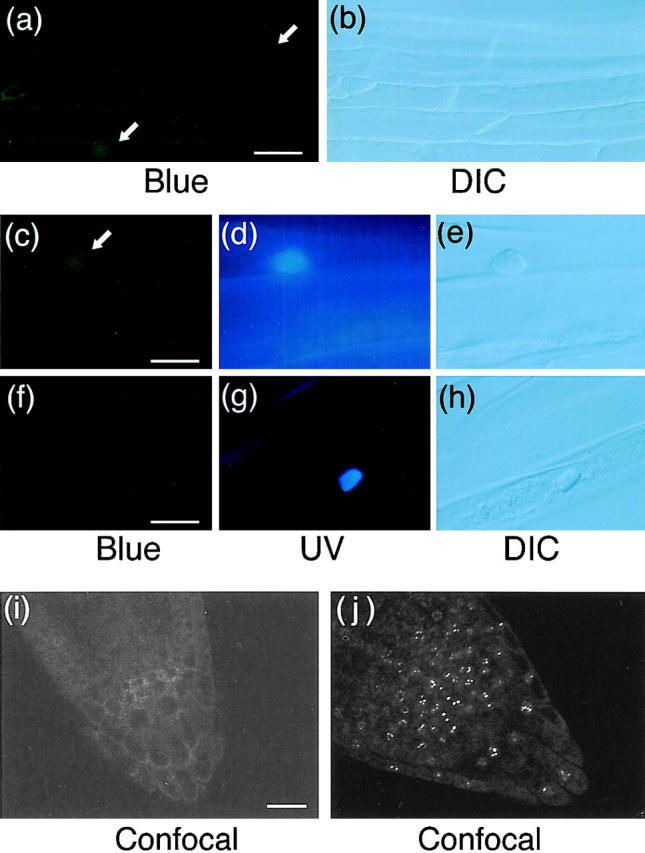
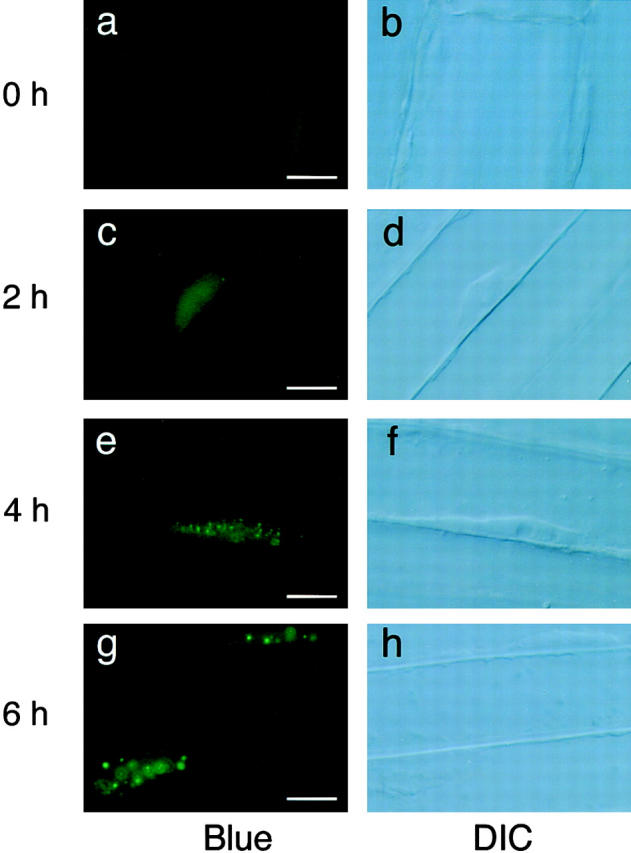
Phytochrome is a ubiquitous photoreceptor of plants and is encoded by a small multigene family. We have shown recently that a functional nuclear localization signal may reside within the COOH-terminal region of a major member of the family, phytochrome B (phyB) (Sakamoto, K., and A. Nagatani. 1996. Plant J. 10:859-868). In the present study, a fusion protein consisting of full-length phyB and the green fluorescent protein (GFP) was overexpressed in the phyB mutant of Arabidopsis to examine subcellular localization of phyB in intact tissues. The resulting transgenic lines exhibited pleiotropic phenotypes reported previously for phyB overexpressing plants, suggesting that the fusion protein is biologically active. Immunoblot analysis with anti-phyB and anti-GFP monoclonal antibodies confirmed that the fusion protein accumulated to high levels in these lines. Fluorescence microscopy of the seedlings revealed that the phyB-GFP fusion protein was localized to the nucleus in light grown tissues. Interestingly, the fusion protein formed speckles in the nucleus. Analysis of confocal optical sections confirmed that the speckles were distributed within the nucleus. In contrast, phyB-GFP fluorescence was observed throughout the cell in dark-grown seedlings. Therefore, phyB translocates to specific sites within the nucleus upon photoreceptor activation.
Figures

References
-
- Ang L-H, Chattopadhyay S, Wei N, Oyama T, Okada K, Batschauer A, Deng X-W. Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsisdevelopment. Mol Cell. 1998;1:213–222. - PubMed
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thalianaplants. CR Acad Sci Paris Life Sci. 1993;316:1194–1199.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
