

Tissue inhibitor of metalloproteinase-2 stimulates mesenchymal growth and regulates epithelial branching during morphogenesis of the rat metanephros
- PMID: 10225973
- PMCID: PMC408351
- DOI: 10.1172/JCI4586
Tissue inhibitor of metalloproteinase-2 stimulates mesenchymal growth and regulates epithelial branching during morphogenesis of the rat metanephros
Abstract
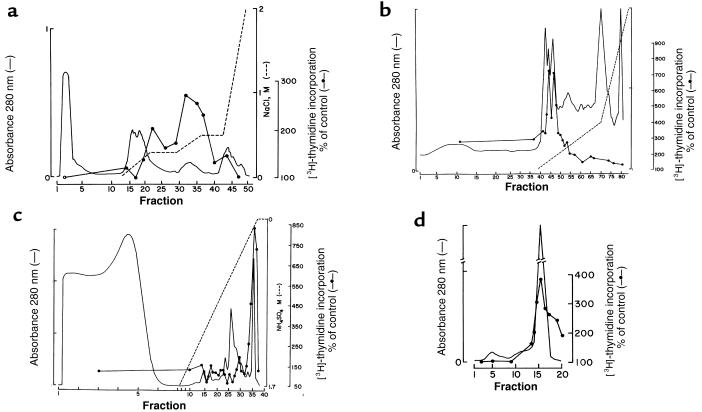
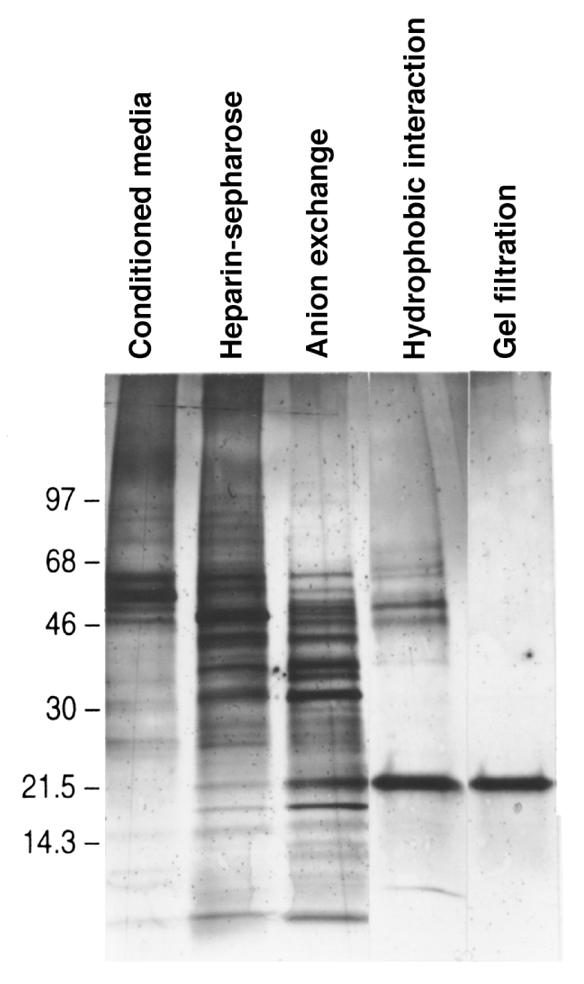
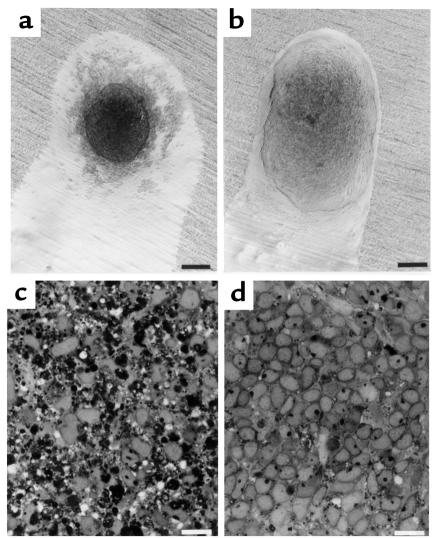
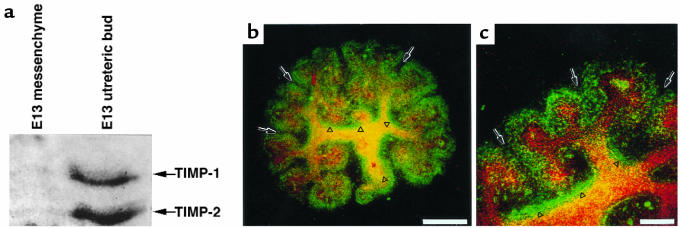
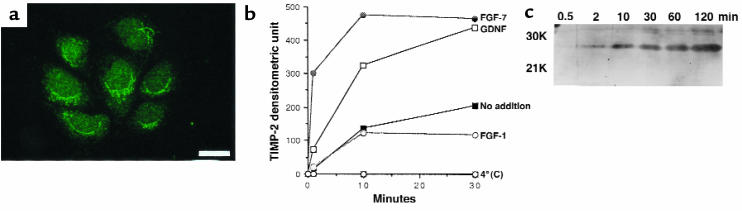
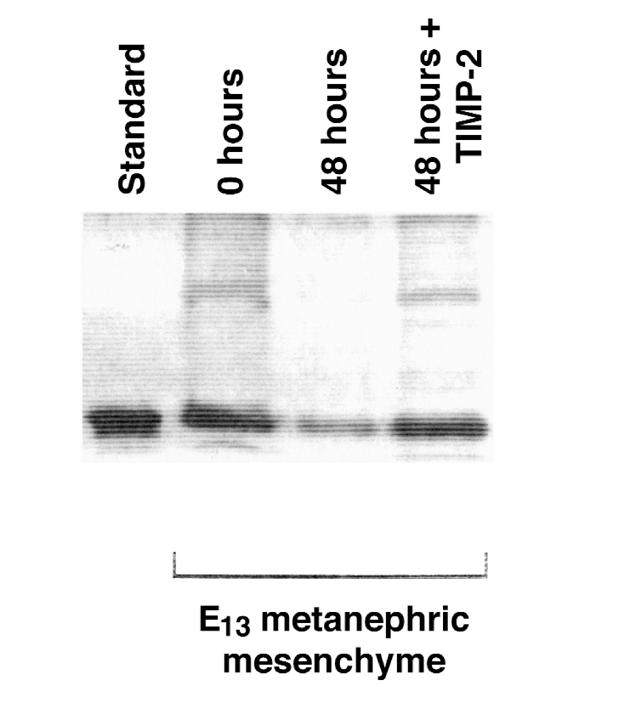
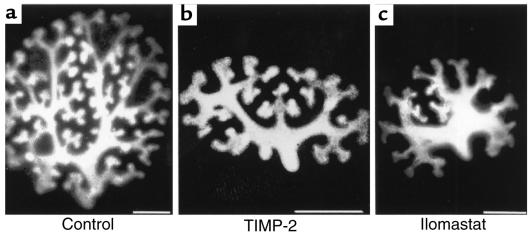
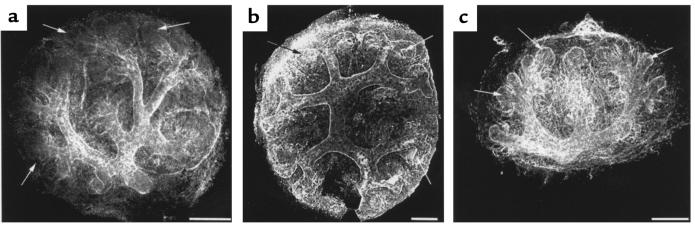
Development of the embryonic kidney results from reciprocal signaling between the ureteric bud and the metanephric mesenchyme. To identify the signaling molecules, we developed an assay in which metanephric mesenchymes are rescued from apoptosis by factors secreted from ureteric bud cells (UB cells). Purification and sequencing of one such factor identified the tissue inhibitor of metalloproteinase-2 (TIMP-2) as a metanephric mesenchymal growth factor. Growth activity was unlikely due to TIMP-2 inhibition of matrix metalloproteinases because ilomastat, a synthetic inhibitor of these enzymes, had no mesenchymal growth action. TIMP-2 was also involved in morphogenesis of the ureteric bud, inhibiting its branching and changing the deposition of its basement membrane; these effects were due to TIMP-2 inhibition of matrix metalloproteinases, as they were reproduced by ilomastat. Thus, TIMP-2 regulates kidney development by at least 2 distinct mechanisms. In addition, TIMP-2 was secreted from UB cells by mesenchymal factors that are essential for ureteric bud development. Hence, the mesenchyme synchronizes its own growth with ureteric morphogenesis by stimulating the secretion of TIMP-2 from the ureteric bud.
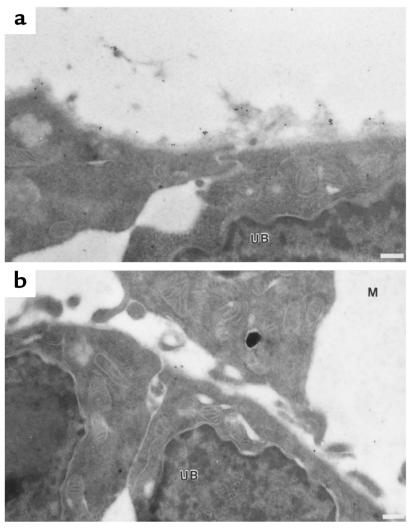
Figures
References
-
- Kreidberg JA, et al. WT-1 is required for early kidney development. Cell. 1993;74:679–691. - PubMed
-
- Sanchez M, et al. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 1996;382:70–73. - PubMed
-
- Pichel J, et al. Defects in enteric innervation and kidney development in mice lacking GDNF. Nature. 1996;382:73–76. - PubMed
-
- Moore M, et al. Renal and neuronal abnormalities in mice lacking GDNF. Nature. 1996;382:76–79. - PubMed
-
- Schuchardt A, et al. Defects in the kidney and the enteric nervous system of mice lacking the tyrosine kinase receptor ret. Nature. 1994;367:380–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
