

Segmentation by color influences responses of motion-sensitive neurons in the cortical middle temporal visual area
- PMID: 10234024
- PMCID: PMC6782728
- DOI: 10.1523/JNEUROSCI.19-10-03935.1999
Segmentation by color influences responses of motion-sensitive neurons in the cortical middle temporal visual area
Abstract
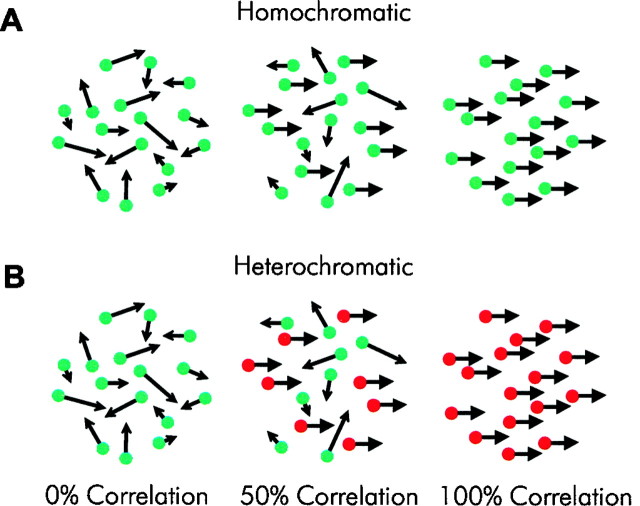
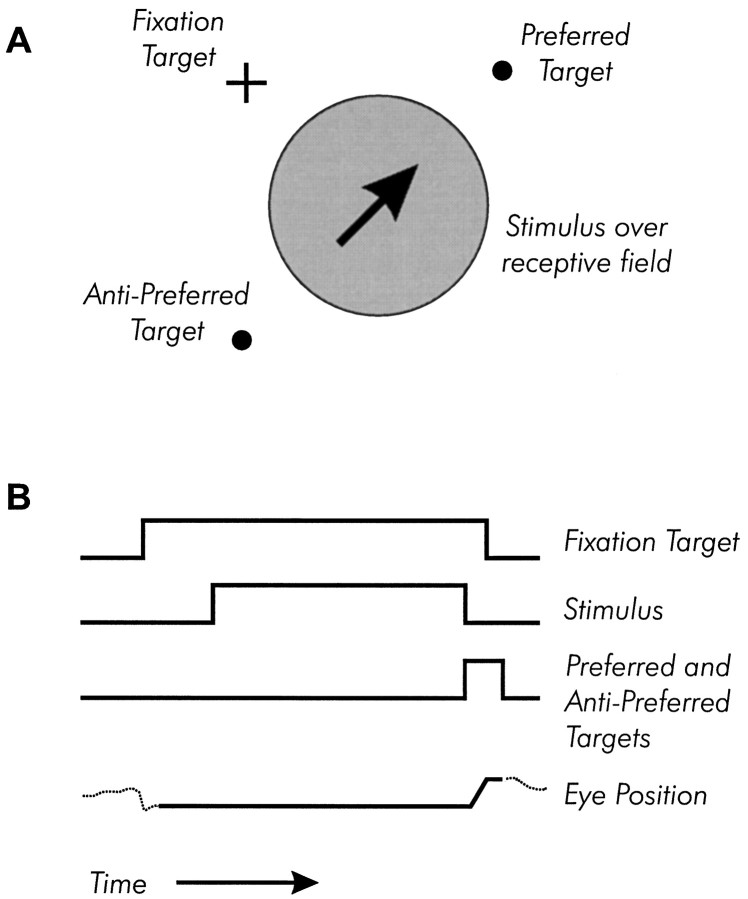
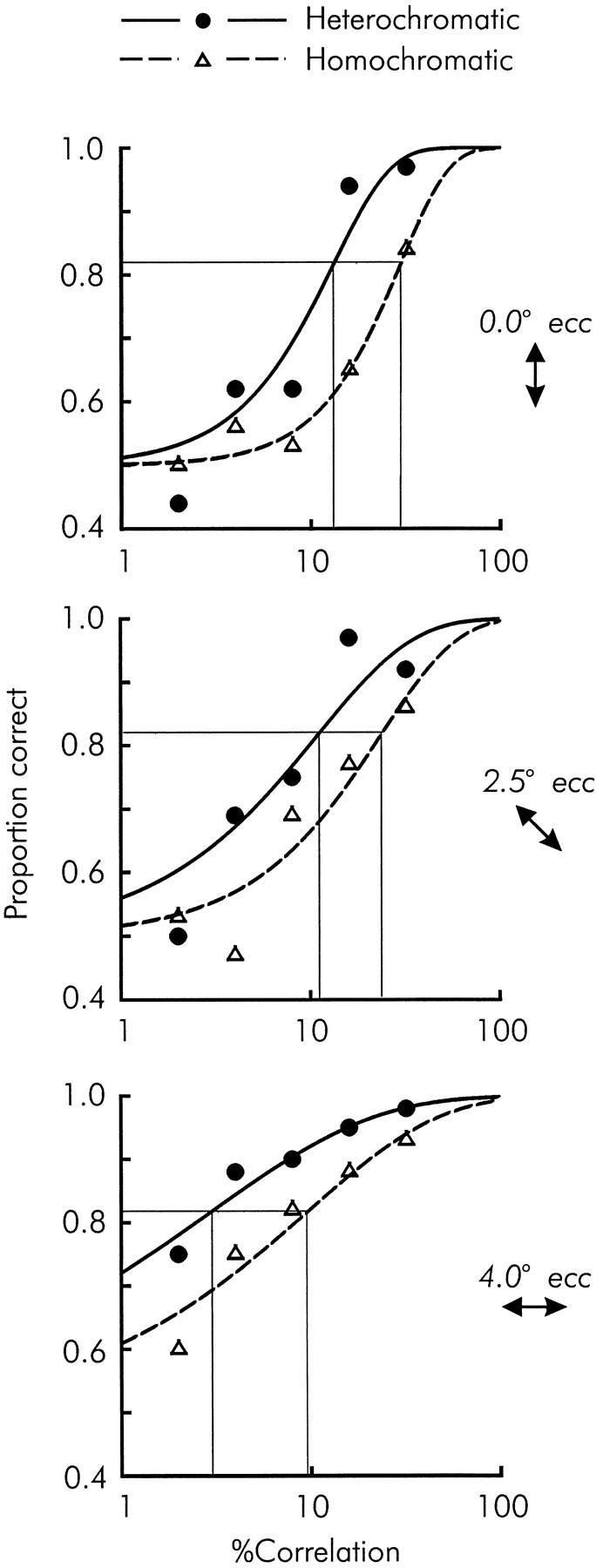
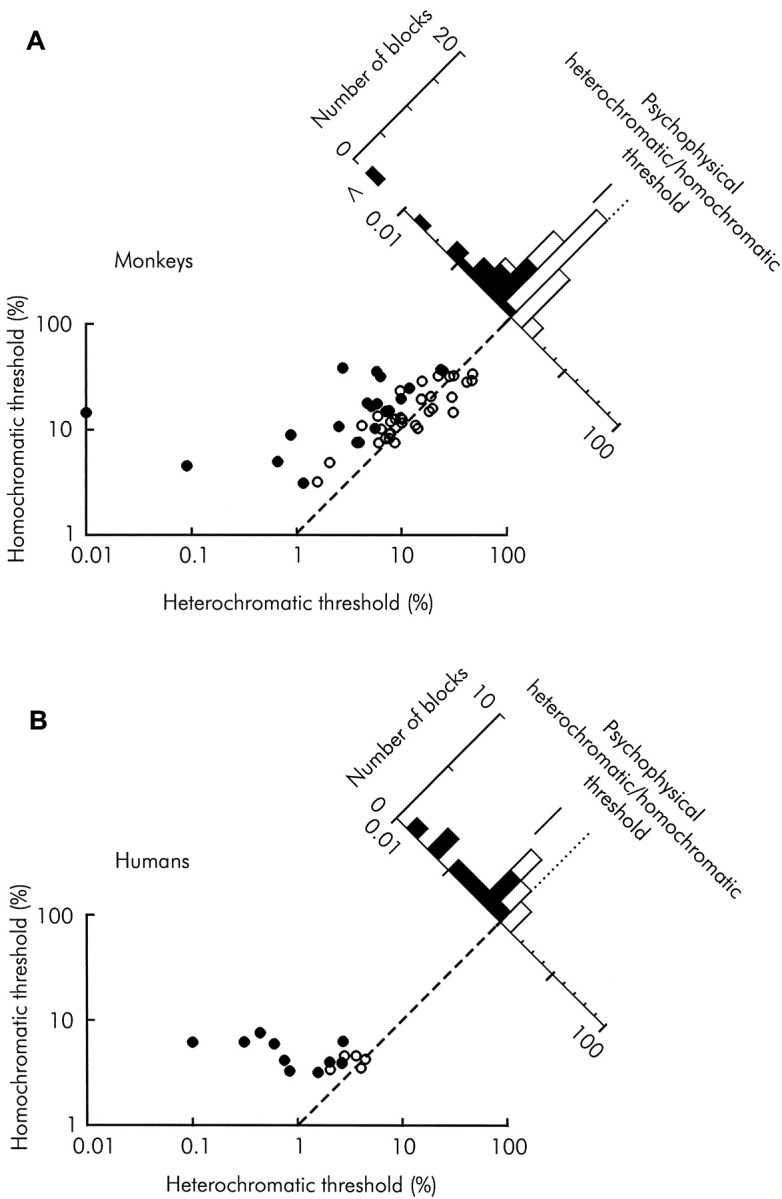
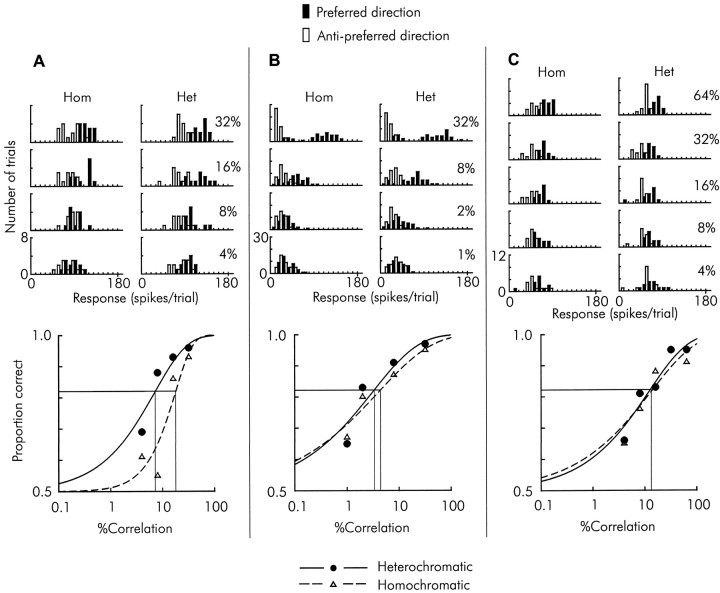
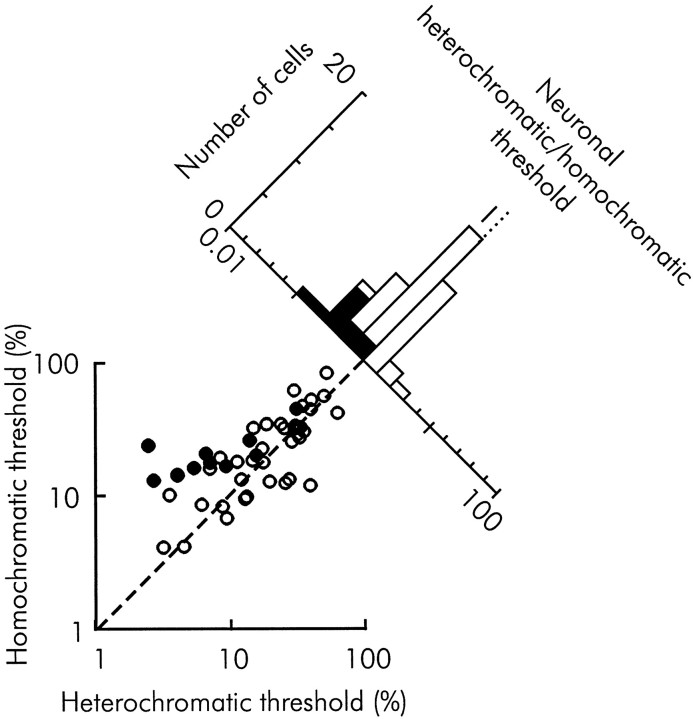
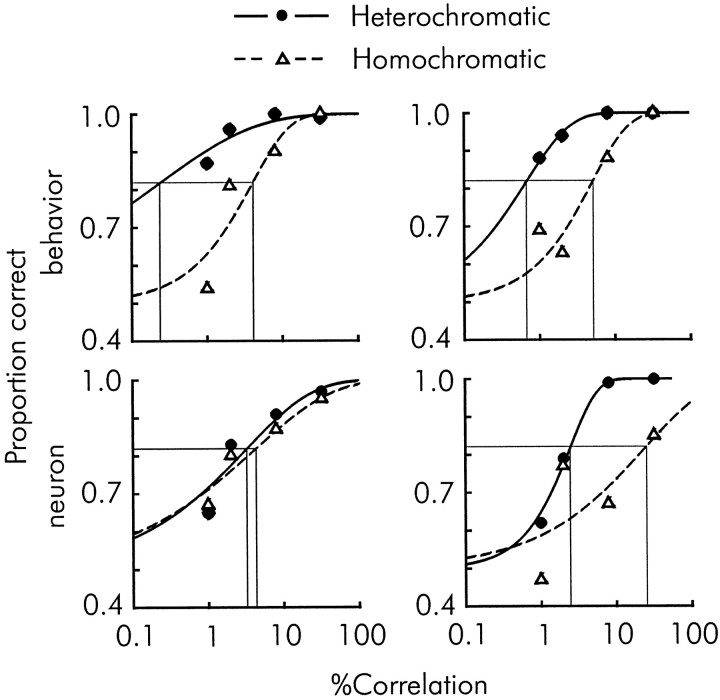
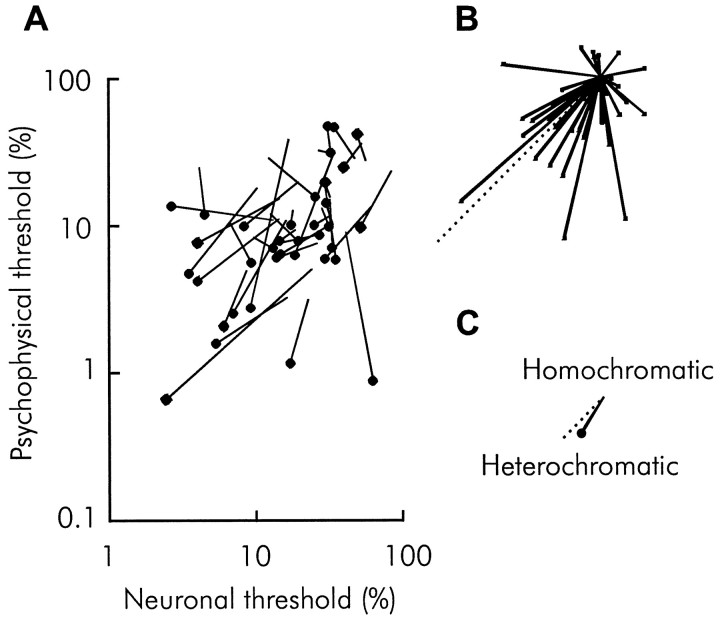
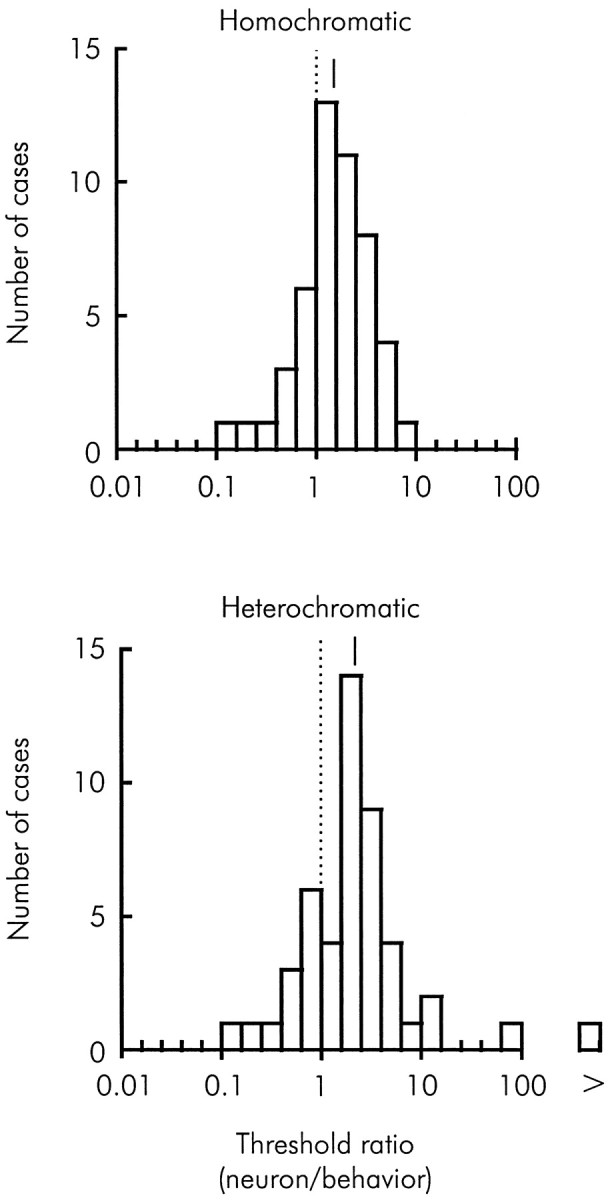
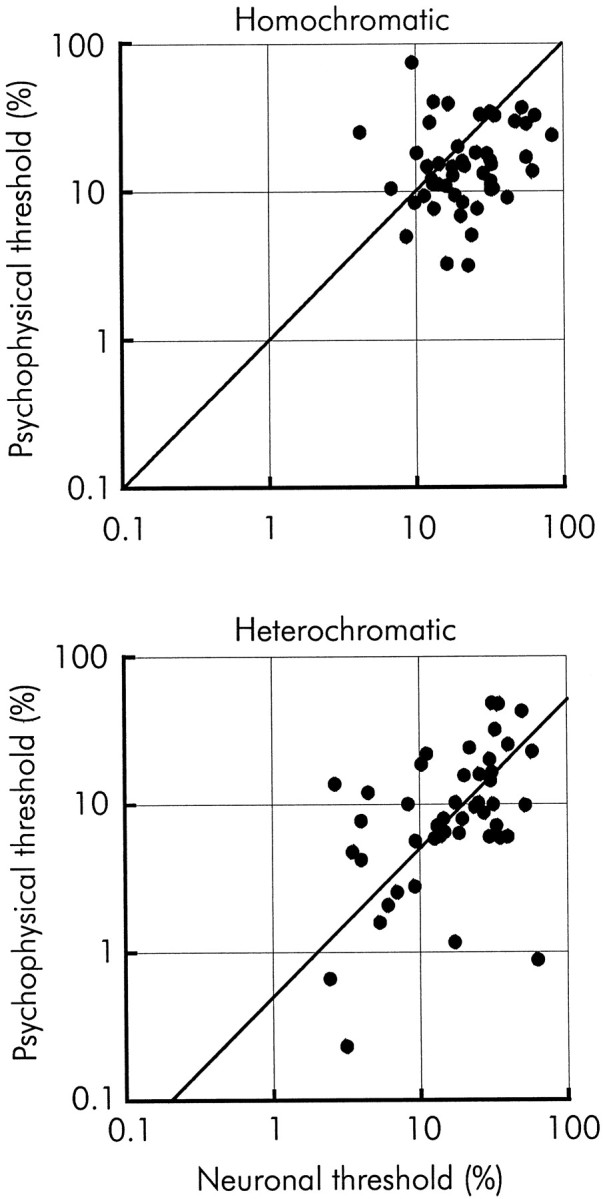
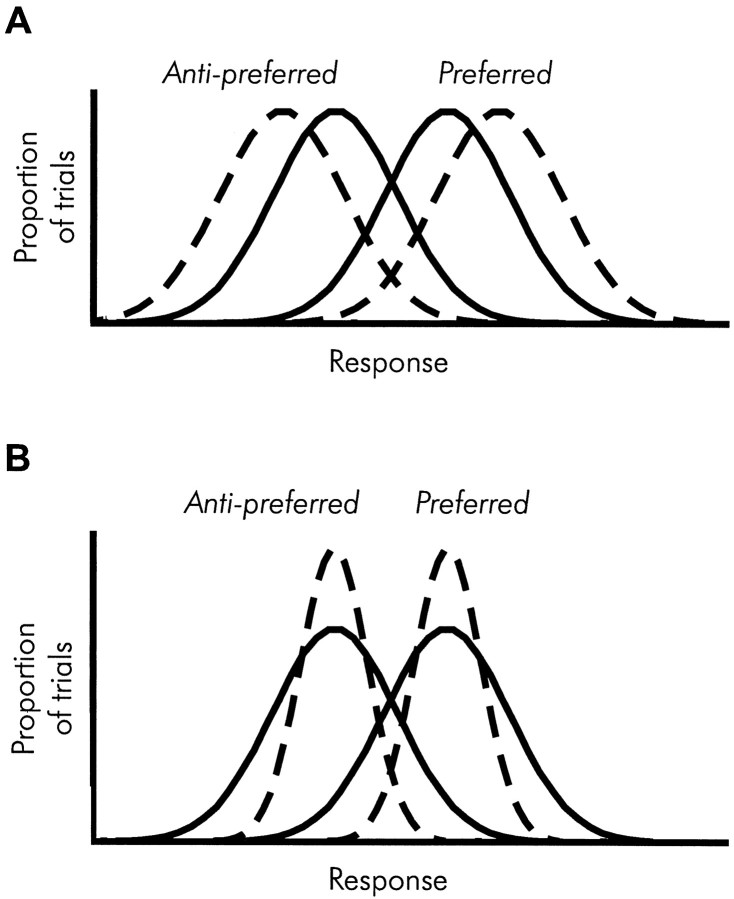
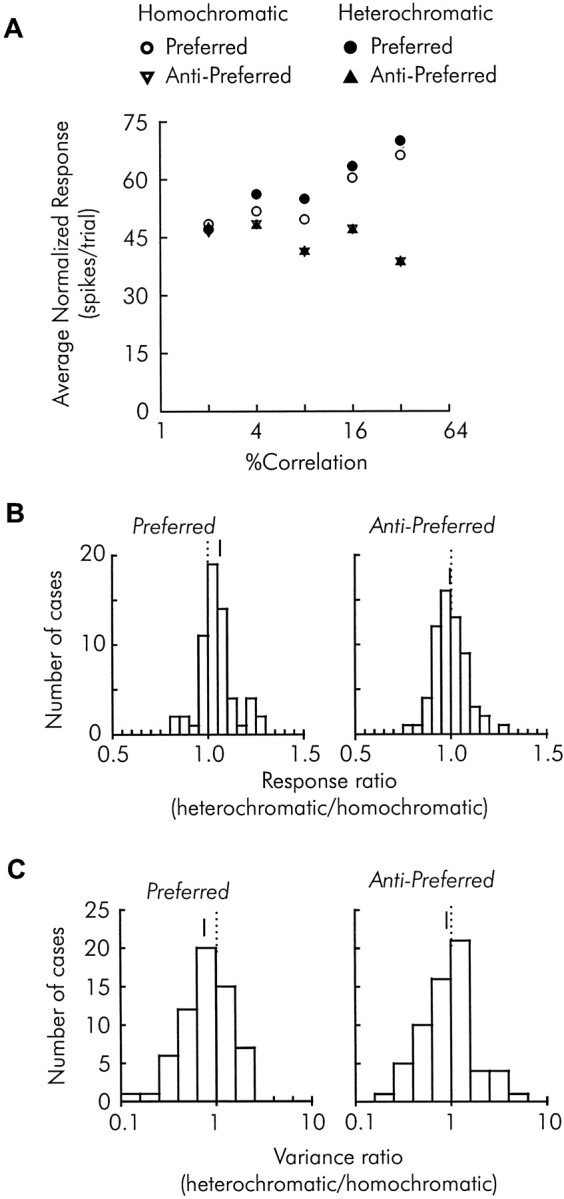
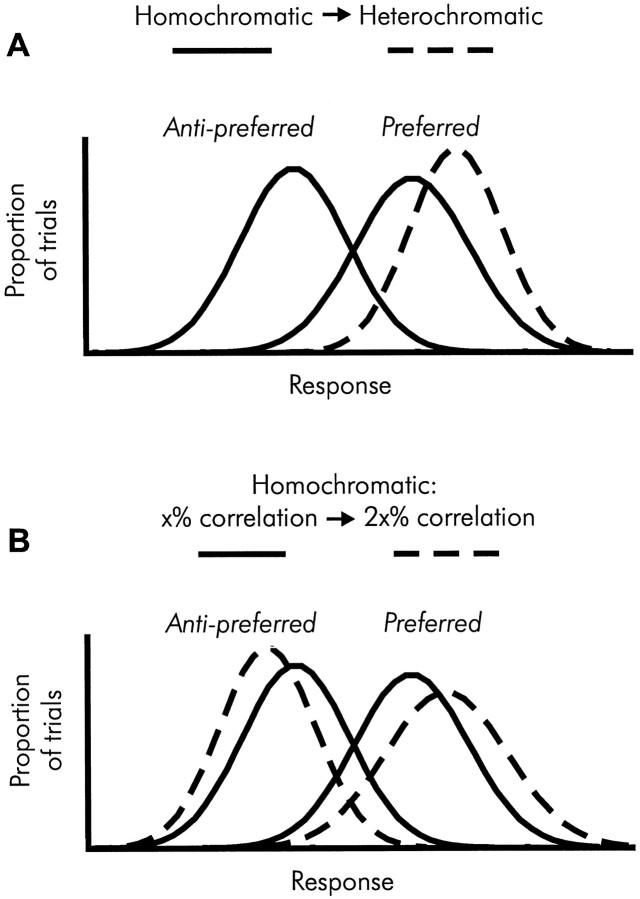
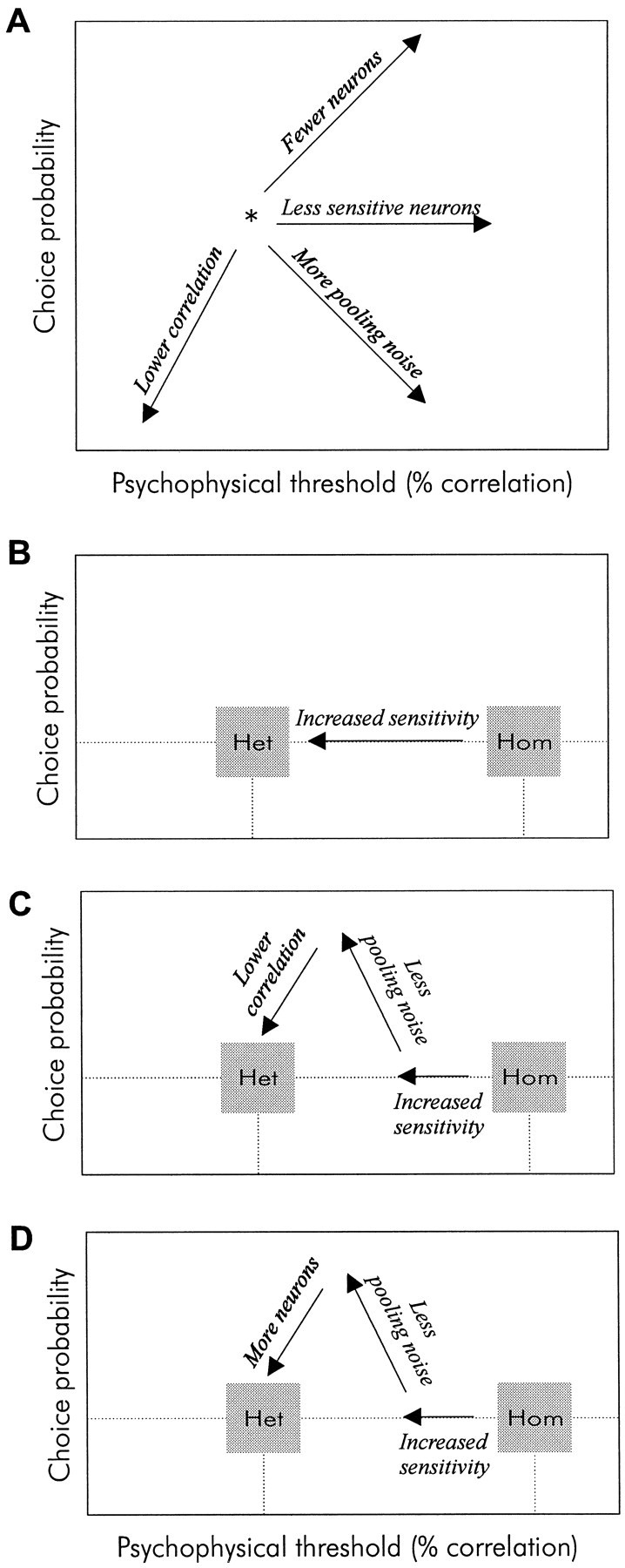
We previously showed that human subjects are better able to discriminate the direction of a motion signal in dynamic noise when the signal is distinguished (segmented) from the noise by color. This finding suggested a hitherto unexplored avenue of interaction between motion and color pathways in the primate visual system. To examine whether chromatic segmentation exerts a similar influence on cortical neurons that contribute to motion direction discrimination, we have now compared the discriminative capacity of single MT neurons and psychophysical observers viewing motion signals with and without chromatic segmentation. All data were obtained from rhesus monkeys trained to discriminate motion direction in dynamic stimuli containing varying proportions of coherently moving (signal) and randomly moving (noise) dots. We obtained psychophysical and neurophysiological data in the same animals, on the same trials, and using the same visual display. Chromatic segmentation of the signal from the noise enhanced both neuronal and psychophysical sensitivity to the motion signal but had a smaller influence on neuronal than on psychophysical sensitivity. Hence the ratio of neuronal to psychophysical thresholds, one measure of the relation between neuronal and psychophysical performance, depended on chromatic segmentation. Increased neuronal sensitivity to chromatically segmented displays stemmed from larger and less noisy responses to motion in the preferred directions of the neurons, suggesting that specialized mechanisms influence responses in the motion pathway when color segments motion signal in visual scenes. These findings lead us to reevaluate potential mechanisms for pooling of MT responses and the role of MT in motion perception.
Figures

Similar articles
-
What happens if it changes color when it moves?: the nature of chromatic input to macaque visual area MT.J Neurosci. 1994 Aug;14(8):4854-70. doi: 10.1523/JNEUROSCI.14-08-04854.1994. J Neurosci. 1994. PMID: 8046456 Free PMC article.
-
Neuronal and psychophysical sensitivity to motion signals in extrastriate area MST of the macaque monkey.J Neurosci. 1994 Jul;14(7):4109-24. doi: 10.1523/JNEUROSCI.14-07-04109.1994. J Neurosci. 1994. PMID: 8027765 Free PMC article.
-
The analysis of visual motion: a comparison of neuronal and psychophysical performance.J Neurosci. 1992 Dec;12(12):4745-65. doi: 10.1523/JNEUROSCI.12-12-04745.1992. J Neurosci. 1992. PMID: 1464765 Free PMC article.
-
Linking Neuronal Direction Selectivity to Perceptual Decisions About Visual Motion.Annu Rev Vis Sci. 2020 Sep 15;6:335-362. doi: 10.1146/annurev-vision-121219-081816. Annu Rev Vis Sci. 2020. PMID: 32936737 Review.
-
Higher cortical visual function.Curr Opin Ophthalmol. 1998 Dec;9(6):40-5. doi: 10.1097/00055735-199812000-00007. Curr Opin Ophthalmol. 1998. PMID: 10387335 Review.
Cited by
-
Neuronal representations of stimulus associations develop in the temporal lobe during learning.Proc Natl Acad Sci U S A. 2001 Oct 9;98(21):12239-44. doi: 10.1073/pnas.211431098. Epub 2001 Sep 25. Proc Natl Acad Sci U S A. 2001. PMID: 11572946 Free PMC article.
-
Attention to the Color of a Moving Stimulus Modulates Motion-Signal Processing in Macaque Area MT: Evidence for a Unified Attentional System.Front Syst Neurosci. 2009 Oct 30;3:12. doi: 10.3389/neuro.06.012.2009. eCollection 2009. Front Syst Neurosci. 2009. PMID: 19893762 Free PMC article.
-
Linking neuronal and behavioral performance in a reaction-time visual detection task.J Neurosci. 2007 Jul 25;27(30):8122-37. doi: 10.1523/JNEUROSCI.1940-07.2007. J Neurosci. 2007. PMID: 17652603 Free PMC article.
-
What constitutes an efficient reference frame for vision?Nat Neurosci. 2002 Oct;5(10):1010-5. doi: 10.1038/nn914. Nat Neurosci. 2002. PMID: 12219092 Free PMC article.
-
Feature binding in the feedback layers of area V2.Cereb Cortex. 2009 Oct;19(10):2230-9. doi: 10.1093/cercor/bhn243. Epub 2009 Jan 19. Cereb Cortex. 2009. PMID: 19153106 Free PMC article.
References
-
- Abramov I, Gordon J, Chan H. Color appearance in the peripheral retina: effects of stimulus size. J Opt Soc Am A. 1991;8:404–414. - PubMed
-
- Britten KH, Shadlen MN, Newsome WT, Movshon JA. Responses of neurons in macaque MT to stochastic motion signals. Vis Neurosci. 1993;10:1157–1169. - PubMed
-
- Britten KH, Newsome WT, Shadlen MN, Celebrini S, Movshon JA. A relationship between behavioral choice and the visual responses of neurons in macaque MT. Vis Neurosci. 1996;13:87–100. - PubMed
-
- Buračas GT, Albright TD. Neural correlates of target detection during visual search in area MT. Soc Neurosci Abstr. 1995;21:1759.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources