

Contributions of the brain angiotensin IV-AT4 receptor subtype system to spatial learning
- PMID: 10234025
- PMCID: PMC6782731
- DOI: 10.1523/JNEUROSCI.19-10-03952.1999
Contributions of the brain angiotensin IV-AT4 receptor subtype system to spatial learning
Abstract
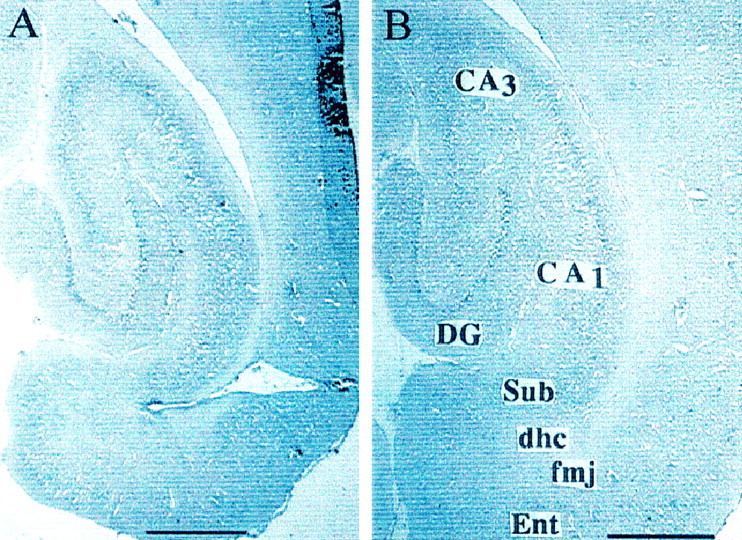
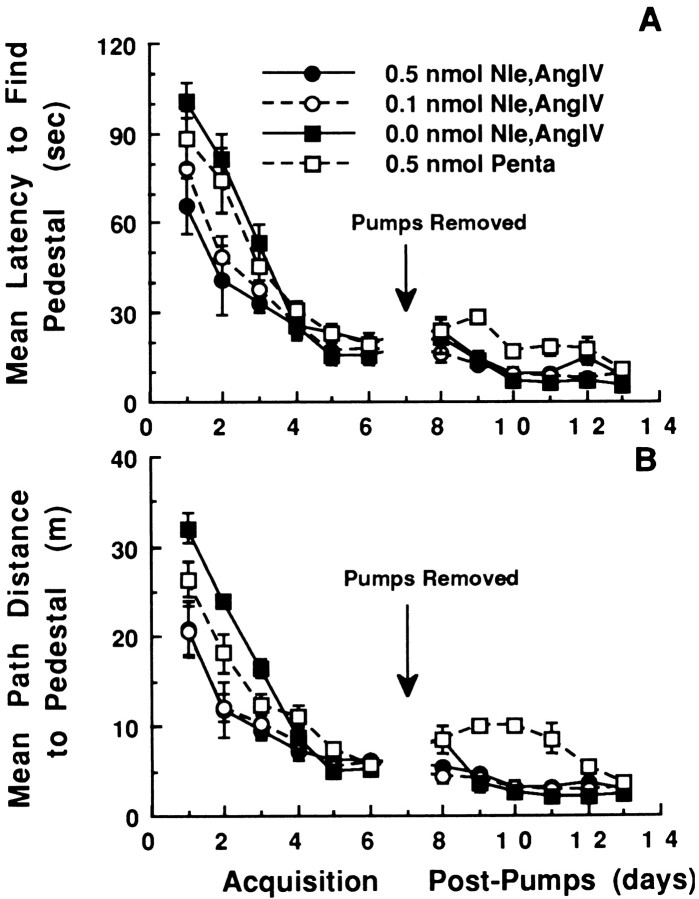
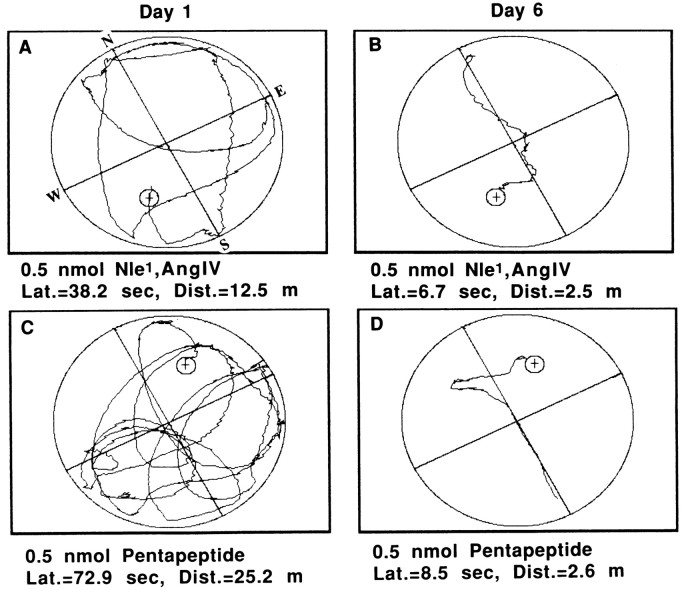
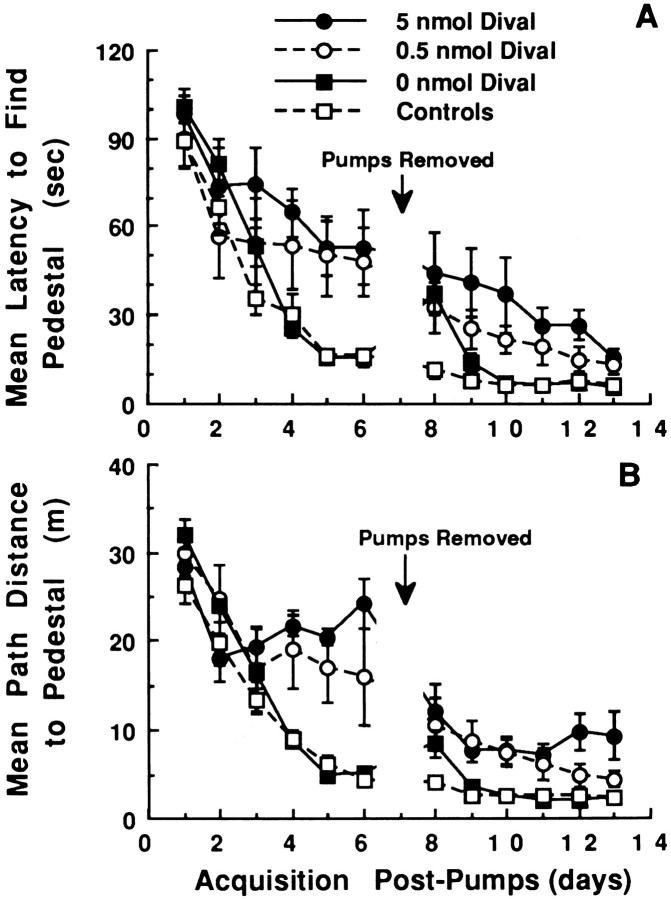
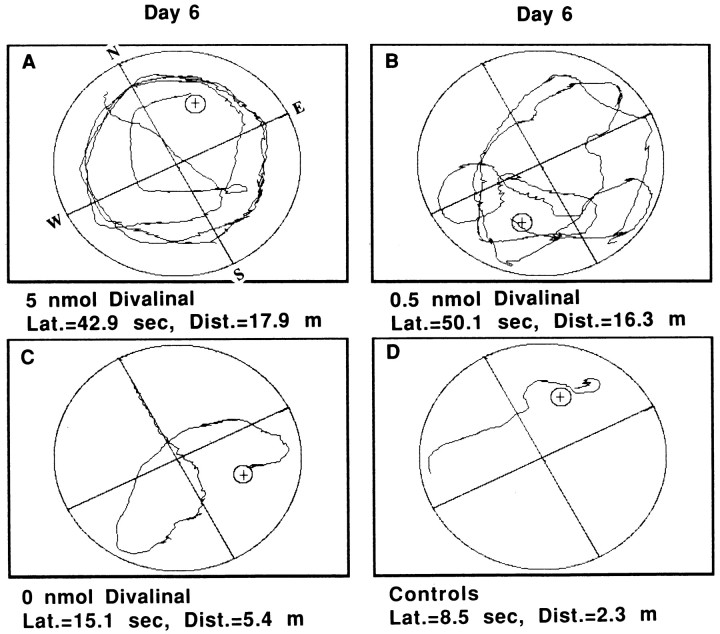
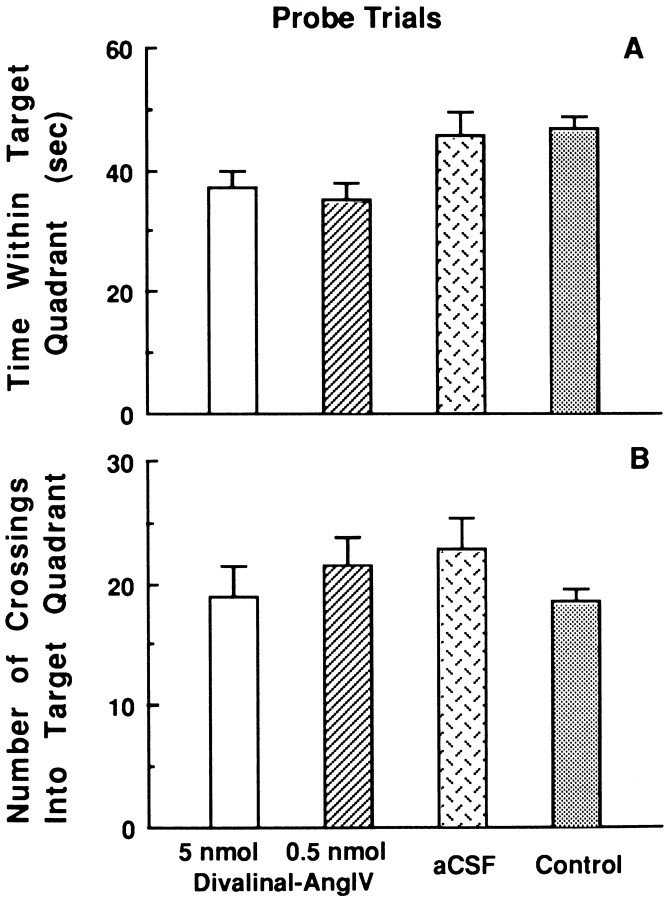
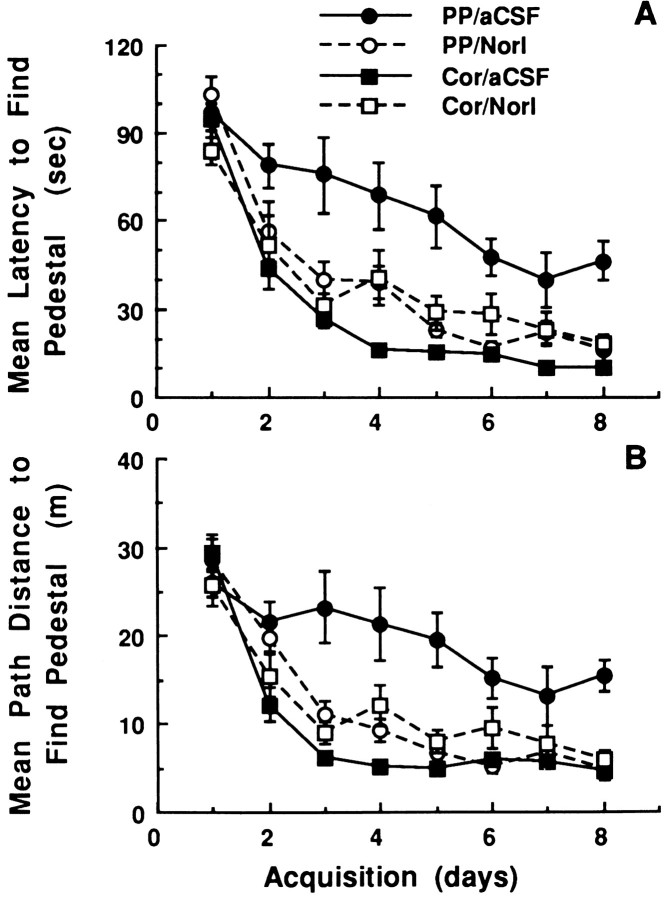
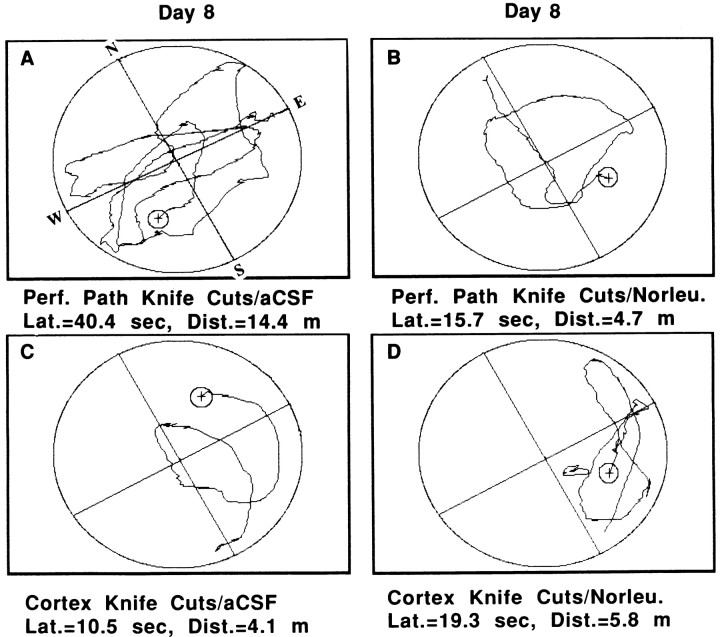
The development of navigational strategies to solve spatial problems appears to be dependent on an intact hippocampal formation. The circular water maze task requires the animal to use extramaze spatial cues to locate a pedestal positioned just below the surface of the water. Presently, we investigated the role of a recently discovered brain angiotensin receptor subtype (AT4) in the acquisition of this spatial learning task. The AT4 receptor subtype is activated by angiotensin IV (AngIV) rather than angiotensins II or III, as documented for the AT1 and AT2 receptor subtypes, and is heavily distributed in the CA1-CA3 fields of the hippocampus. Chronic intracerebroventricular infusion of a newly synthesized AT4 agonist (Norleucine1-AngIV) via osmotic pump facilitated the rate of acquisition to solve this task, whereas treatment with an AT4 receptor antagonist (Divalinal) significantly interfered with the acquisition of successful search strategies. Animals prepared with bilateral knife cuts of the perforant path, a major afferent hippocampal fiber bundle originating in the entorhinal cortex, displayed deficits in solving this task. This performance deficit could be reversed with acute intracerebroventricular infusion of a second AT4 receptor agonist (Norleucinal). These results suggest that the brain AngIV-AT4 system plays a role in the formation of spatial search strategies and memories. Further, application of an AT4 receptor agonist compensated for spatial memory deficits in performance accompanying perforant path knife cuts. Possible mechanisms underlying this compensatory effect are discussed.
Figures
References
-
- Agnihotri N, Lopez-Garcia JC, Hawkins RD, Arancio O. Morphological changes associated with long-term potentiation. Histol Histopathol. 1998;13:1155–1162. - PubMed
-
- Albrecht D, Broser M, Kruger H. Excitatory action of angiotensins II and IV on hippocampal neuronal activity in urethane anesthetized rats. Regul Pept. 1997a;70:105–109. - PubMed
-
- Albrecht D, Broser M, Kruger H, Bader M. Effects of angiotensin II and IV on geniculate activity in nontransgenic and transgenic rats. Eur J Pharmacol. 1997b;332:53–63. - PubMed
-
- Braszko JJ, Kupryszewski G, Witczuk B, Wisniewski K. Angiotensin II-(3–8)-hexapeptide affects motor activity, performance of passive avoidance and a conditioned avoidance response in rats. Neuroscience. 1988;27:777–783. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous