

The stimulatory action and the development of tolerance to caffeine is associated with alterations in gene expression in specific brain regions
- PMID: 10234030
- PMCID: PMC6782739
- DOI: 10.1523/JNEUROSCI.19-10-04011.1999
The stimulatory action and the development of tolerance to caffeine is associated with alterations in gene expression in specific brain regions
Abstract
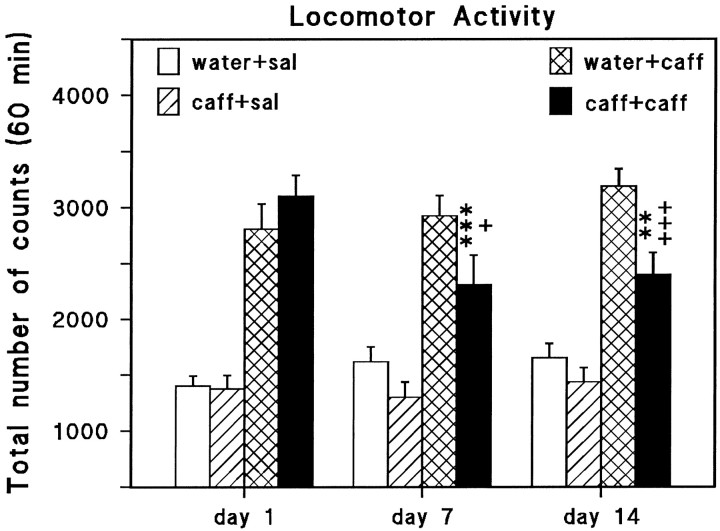
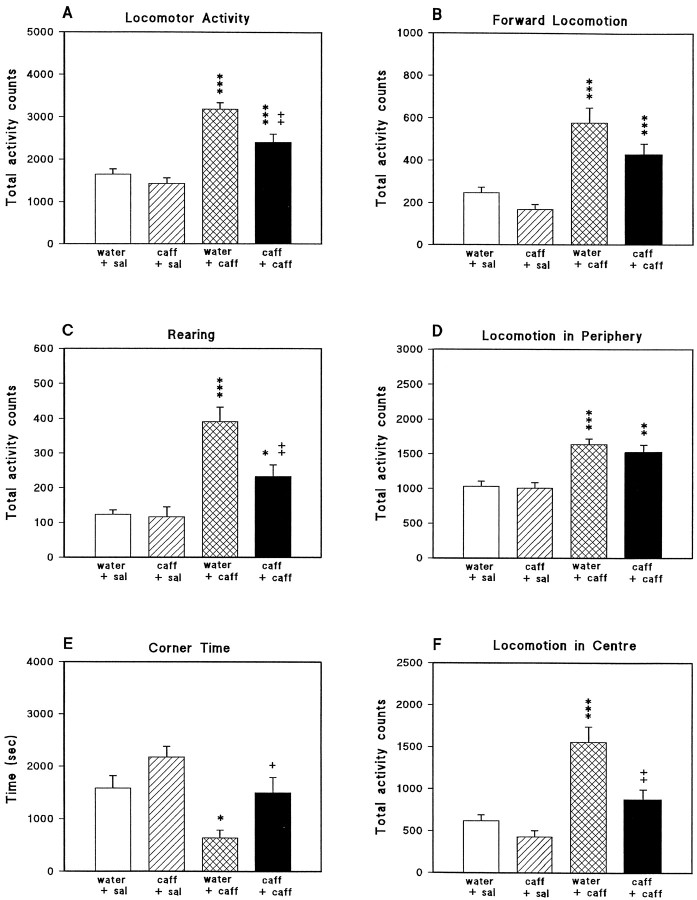
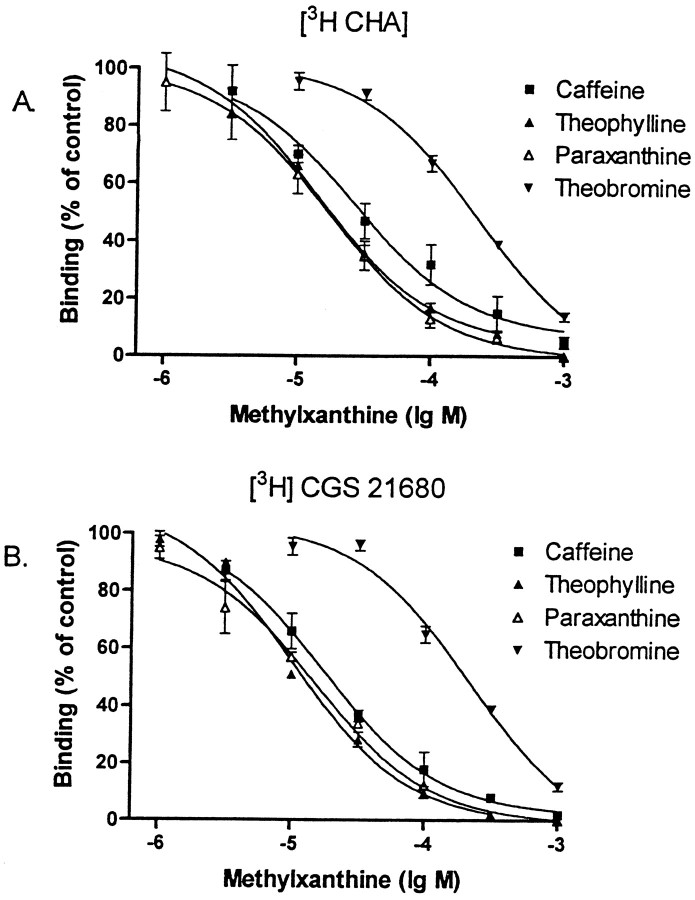
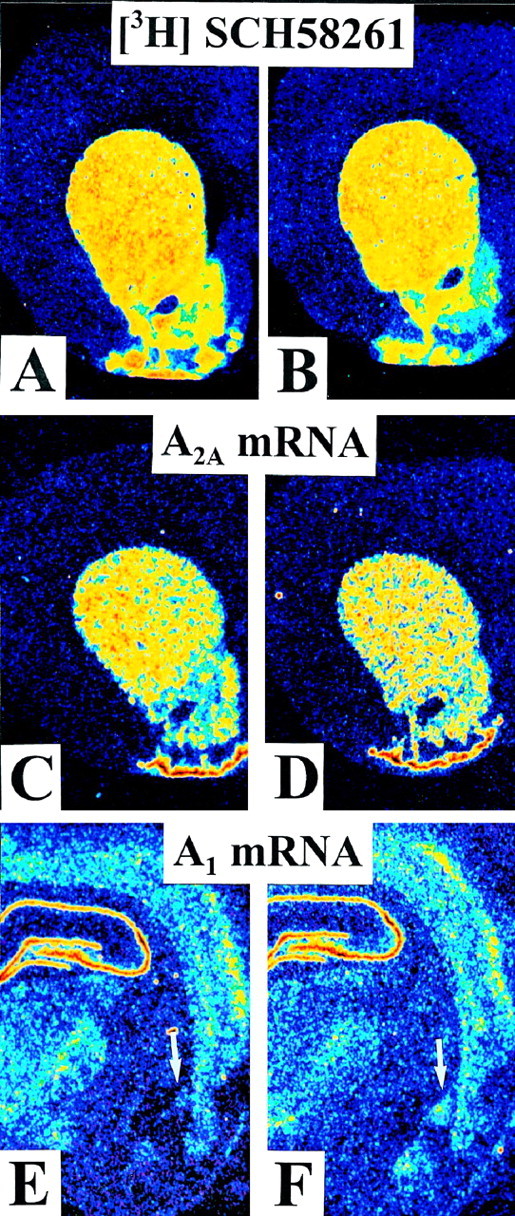
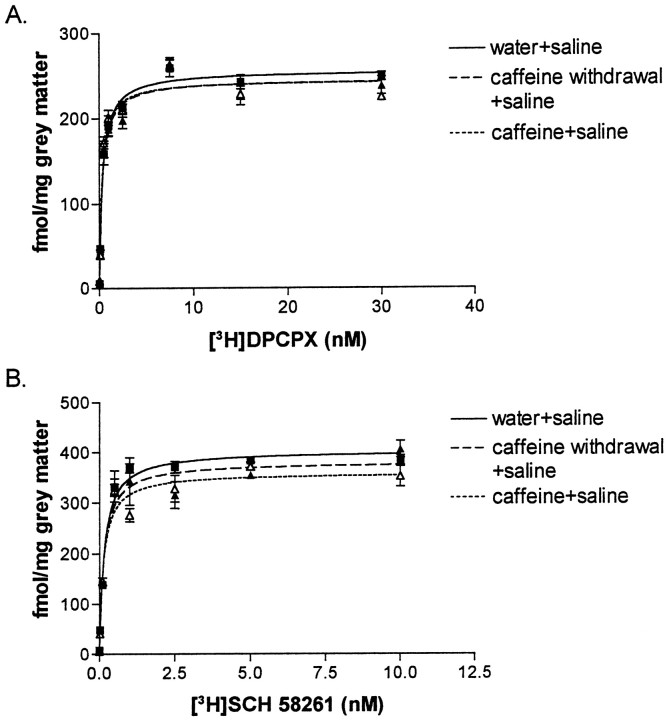
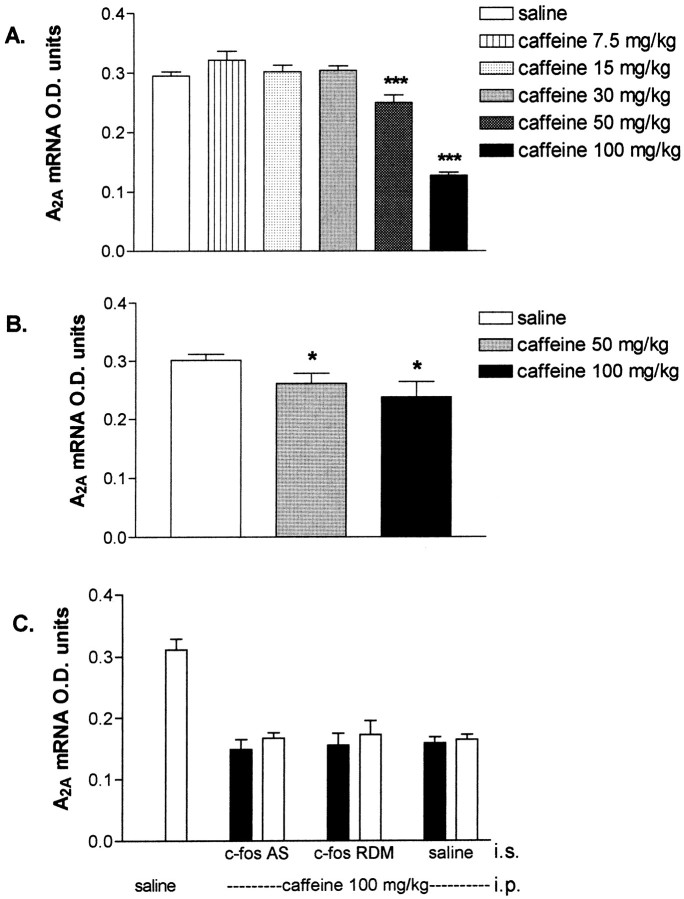
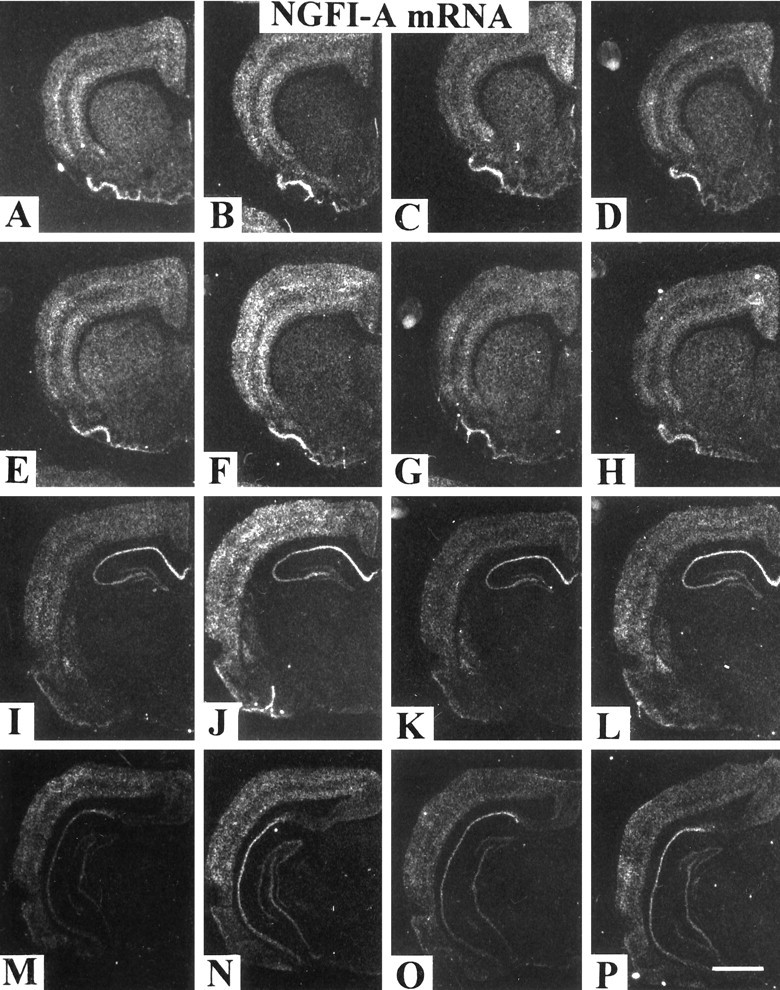
We sought neurochemical correlates to the stimulatory action of caffeine in rats and to adaptations during development of tolerance. Acute intraperitoneal injections of caffeine (7.5 mg/kg) increased locomotion and NGFI-A mRNA, a marker of neuronal activity, in the hippocampal area CA1, but decreased NGFI-A mRNA in rostral striatum and nucleus accumbens. Rats that received caffeine (0.3 gm/l) in their drinking water for 14 d developed tolerance to the stimulatory effect of a challenge with caffeine (7.5 mg/kg) and responded with a less pronounced decrease of NGFI-A mRNA in rostral striatum and nucleus accumbens. Metabolism of caffeine to its active metabolites was increased in tolerant animals, but the total level of active metabolites in brain was not significantly altered. Thus, there are changes in caffeine metabolism after long-term caffeine treatment, but they cannot explain development of tolerance. Caffeine-tolerant animals had downregulated levels of adenosine A2A receptors and the corresponding mRNA in rostral parts of striatum, but an increased expression of adenosine A1 receptor mRNA in the lateral amygdala. No changes in mesencephalic tyrosine hydroxylase mRNA were found in caffeine-tolerant rats. Thus, we have identified neuronal pathways that are regulated by adenosine A1 and/or A2A receptors and are targets for the stimulatory action of caffeine. Furthermore, adaptive changes in gene expression in these brain areas were associated with the development of locomotor tolerance to caffeine.
Figures
References
-
- Arslan G, Kull B, Fredholm BB. Signaling via A2A adenosine receptors in four PC 12 cell clones. Naunyn Schmiedebergs Arch Pharmacol. 1999;359:28–32. - PubMed
-
- Barraco RA, Martens KA, Parizon M, Normile HJ. Adenosine A2a receptors in the nucleus accumbens mediate locomotor depression. Brain Res Bull. 1993;31:397–404. - PubMed
-
- Beitner-Johnson D, Nestler EJ. Morphine and cocaine exert common chronic actions on tyrosine hydroxylase in dopaminergic brain reward regions. J Neurochem. 1991;57:344–347. - PubMed
-
- Boissier JR, Simon P. Action de la cafeine sur la motilite spontanee de la souris. Arch Int Pharmacodyn Ther. 1965;158:212–221. - PubMed
-
- Brockwell NT, Eikelboom R, Beninger RJ. Caffeine-induced place and taste conditioning: production of dose-dependent preference and aversion. Pharmacol Biochem Behav. 1991;38:513–517. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous