

Arabidopsis 22-kilodalton peroxisomal membrane protein. Nucleotide sequence analysis and biochemical characterization
- PMID: 10318708
- PMCID: PMC59263
- DOI: 10.1104/pp.120.1.309
Arabidopsis 22-kilodalton peroxisomal membrane protein. Nucleotide sequence analysis and biochemical characterization
Abstract
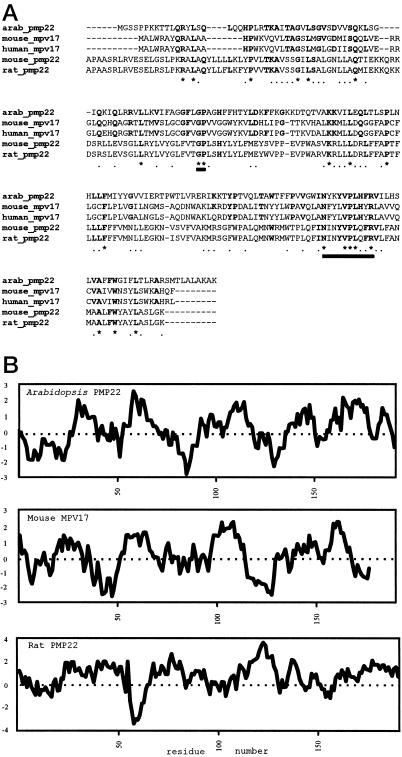
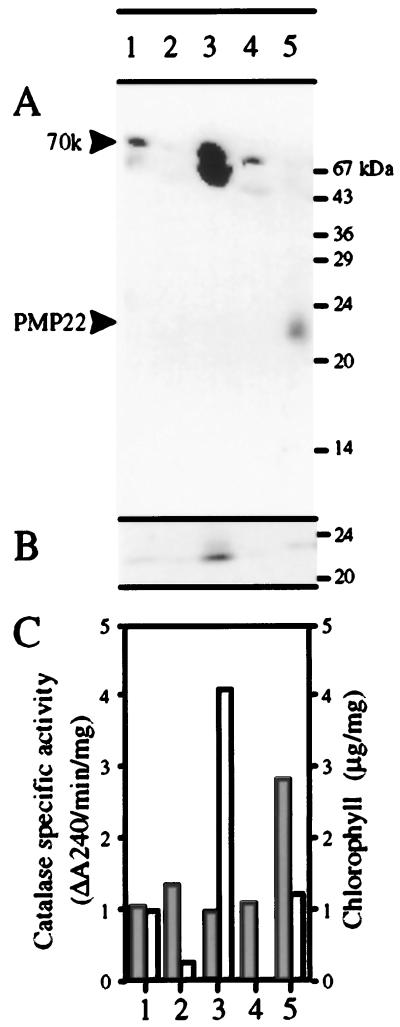
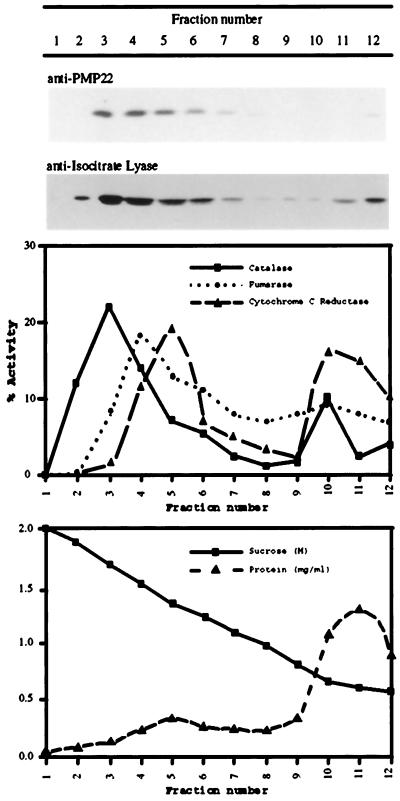
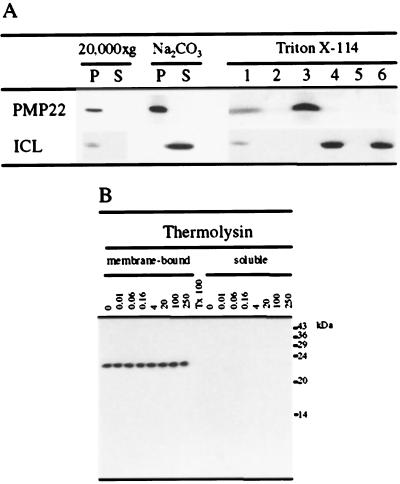
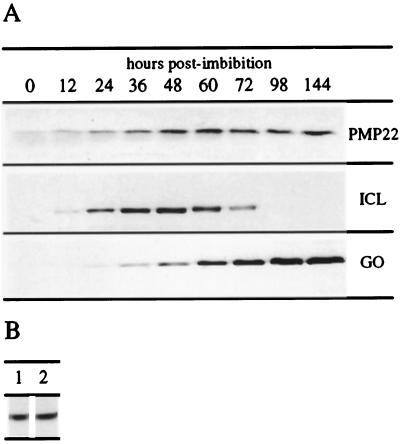
We sequenced and characterized PMP22 (22-kD peroxisomal membrane protein) from Arabidopsis, which shares 28% to 30% amino acid identity and 55% to 57% similarity to two related mammalian peroxisomal membrane proteins, PMP22 and Mpv17. Subcellular fractionation studies confirmed that the Arabidopsis PMP22 is a genuine peroxisomal membrane protein. Biochemical analyses established that the Arabidopsis PMP22 is an integral membrane protein that is completely embedded in the lipid bilayer. In vitro import assays demonstrated that the protein is inserted into the membrane posttranslationally in the absence of ATP, but that ATP stimulates the assembly into the native state. Arabidopsis PMP22 is expressed in all organs of the mature plant and in tissue-cultured cells. Expression of PMP22 is not associated with a specific peroxisome type, as it is detected in seeds and throughout postgerminative growth as cotyledon peroxisomes undergo conversion from glyoxysomes to leaf-type peroxisomes. Although PMP22 shows increased accumulation during the growth of young seedlings, its expression is not stimulated by light.
Figures

Similar articles
-
Characterization of the targeting signal of the Arabidopsis 22-kD integral peroxisomal membrane protein.Plant Physiol. 2003 Oct;133(2):813-28. doi: 10.1104/pp.103.027870. Epub 2003 Sep 4. Plant Physiol. 2003. PMID: 12972647 Free PMC article.
-
Developmental analysis of a putative ATP/ADP carrier protein localized on glyoxysomal membranes during the peroxisome transition in pumpkin cotyledons.Plant Cell Physiol. 2001 Aug;42(8):835-41. doi: 10.1093/pcp/pce108. Plant Cell Physiol. 2001. PMID: 11522909
-
The Arabidopsis PEX12 gene is required for peroxisome biogenesis and is essential for development.Plant Physiol. 2005 Sep;139(1):231-9. doi: 10.1104/pp.105.066811. Epub 2005 Aug 19. Plant Physiol. 2005. PMID: 16113209 Free PMC article.
-
Proteomic identification and characterization of a novel peroxisomal adenine nucleotide transporter supplying ATP for fatty acid beta-oxidation in soybean and Arabidopsis.Plant Cell. 2008 Dec;20(12):3227-40. doi: 10.1105/tpc.108.062877. Epub 2008 Dec 10. Plant Cell. 2008. PMID: 19073762 Free PMC article.
-
Functional transformation of microbodies in higher plant cells.Cell Struct Funct. 1996 Oct;21(5):387-93. doi: 10.1247/csf.21.387. Cell Struct Funct. 1996. PMID: 9118245 Review.
Cited by
-
SYM1 is the stress-induced Saccharomyces cerevisiae ortholog of the mammalian kidney disease gene Mpv17 and is required for ethanol metabolism and tolerance during heat shock.Eukaryot Cell. 2004 Jun;3(3):620-31. doi: 10.1128/EC.3.3.620-631.2004. Eukaryot Cell. 2004. PMID: 15189984 Free PMC article.
-
Deciphering variations, identification of marker-trait associations and candidate genes for seed oil content under terminal heat stress in Indian mustard (Brassica juncea L. Czern & Coss) germplasm stock.3 Biotech. 2024 May;14(5):140. doi: 10.1007/s13205-024-03985-w. Epub 2024 Apr 28. 3 Biotech. 2024. PMID: 38689736 Free PMC article.
-
Arabidopsis AtMPV17, a homolog of mice MPV17, enhances osmotic stress tolerance.Physiol Mol Biol Plants. 2020 Jul;26(7):1341-1348. doi: 10.1007/s12298-020-00834-x. Epub 2020 Jun 13. Physiol Mol Biol Plants. 2020. PMID: 32647452 Free PMC article.
-
Arabidopsis A BOUT DE SOUFFLE, which is homologous with mammalian carnitine acyl carrier, is required for postembryonic growth in the light.Plant Cell. 2002 Sep;14(9):2161-73. doi: 10.1105/tpc.002485. Plant Cell. 2002. PMID: 12215513 Free PMC article.
-
Peroxicretion: a novel secretion pathway in the eukaryotic cell.BMC Biotechnol. 2009 May 20;9:48. doi: 10.1186/1472-6750-9-48. BMC Biotechnol. 2009. PMID: 19457257 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Behari R, Baker A. The carboxy terminus of isocitrate lyase is not essential for import into glyoxysomes in an in vitro system. J Biol Chem. 1993;268:7315–7322. - PubMed
-
- Bordier C. Phase separation of integral membrane proteins in Triton X-114 solution. J Biol Chem. 1981;256:1604–1607. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
