

Transferrin receptor is negatively modulated by the hemochromatosis protein HFE: implications for cellular iron homeostasis
- PMID: 10318901
- PMCID: PMC21877
- DOI: 10.1073/pnas.96.10.5434
Transferrin receptor is negatively modulated by the hemochromatosis protein HFE: implications for cellular iron homeostasis
Abstract
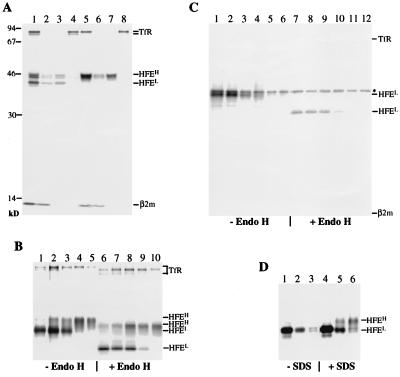
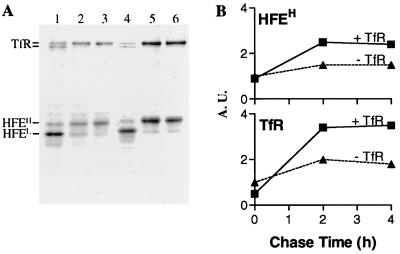
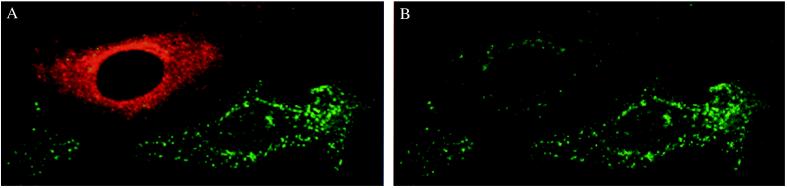
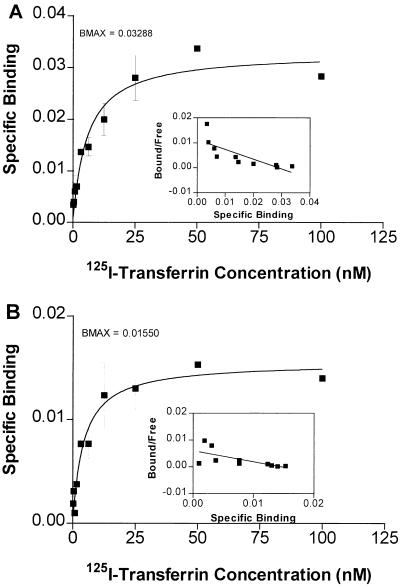
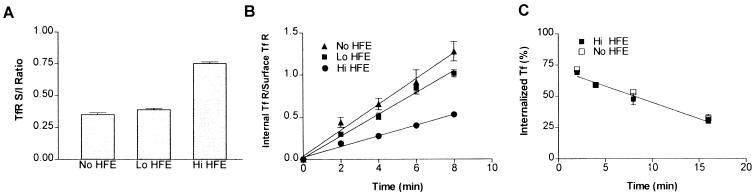
Hereditary hemochromatosis is a common autosomal recessive disorder of iron metabolism. Recent demonstration of an association between transferrin receptor (TfR) and HFE, a major histocompatibility complex class I-like molecule that has been implicated to play a role in hereditary hemochromatosis, further strengthens the notion that HFE is involved in iron metabolism. Herein we show that TfR is required for and controls the assembly and the intracellular transport and surface expression of HFE. Because surface-expressed HFE and TfR remain firmly associated physically, only the fraction of TfR that is associated with HFE during biosynthesis is affected functionally. Moreover, we show that HFE binding reduces the number of functional transferrin binding sites and impairs TfR internalization, thus reducing the uptake of transferrin-bound iron. Thus, iron homeostasis is indirectly regulated by HFE, a negative modulator of TfR.
Figures
Similar articles
-
The major histocompatibility complex-encoded class I-like HFE abrogates endocytosis of transferrin receptor by inducing receptor phosphorylation.Genes Immun. 2000 Oct;1(7):409-17. doi: 10.1038/sj.gene.6363697. Genes Immun. 2000. PMID: 11196670
-
Crystal structure of the hemochromatosis protein HFE and characterization of its interaction with transferrin receptor.Cell. 1998 Apr 3;93(1):111-23. doi: 10.1016/s0092-8674(00)81151-4. Cell. 1998. PMID: 9546397
-
Binding to the transferrin receptor is required for endocytosis of HFE and regulation of iron homeostasis.Nat Cell Biol. 2000 Dec;2(12):953-7. doi: 10.1038/35046611. Nat Cell Biol. 2000. Retraction in: Nat Cell Biol. 2003 Jul;5(7):680. doi: 10.1038/ncb0703-680a. PMID: 11146662 Retracted.
-
Interaction of the hemochromatosis gene product HFE with transferrin receptor modulates cellular iron metabolism.Nutr Rev. 1998 Dec;56(12):356-8. doi: 10.1111/j.1753-4887.1998.tb01679.x. Nutr Rev. 1998. PMID: 9884584 Review.
-
Pumping iron: the strange partnership of the hemochromatosis protein, a class I MHC homolog, with the transferrin receptor.Traffic. 2001 Mar;2(3):167-74. doi: 10.1034/j.1600-0854.2001.020303.x. Traffic. 2001. PMID: 11260522 Review.
Cited by
-
The Q283P amino-acid change in HFE leads to structural and functional consequences similar to those described for the mutated 282Y HFE protein.Hum Genet. 2005 Sep;117(5):467-75. doi: 10.1007/s00439-005-1307-y. Epub 2005 Jun 18. Hum Genet. 2005. PMID: 15965644
-
The hemochromatosis protein HFE inhibits iron export from macrophages.Proc Natl Acad Sci U S A. 2002 Nov 26;99(24):15602-7. doi: 10.1073/pnas.242614699. Epub 2002 Nov 12. Proc Natl Acad Sci U S A. 2002. PMID: 12429850 Free PMC article.
-
Liver iron transport.World J Gastroenterol. 2007 Sep 21;13(35):4725-36. doi: 10.3748/wjg.v13.i35.4725. World J Gastroenterol. 2007. PMID: 17729394 Free PMC article. Review.
-
The haemochromatosis protein HFE induces an apparent iron-deficient phenotype in H1299 cells that is not corrected by co-expression of beta 2-microglobulin.Biochem J. 2003 Mar 15;370(Pt 3):891-9. doi: 10.1042/BJ20021607. Biochem J. 2003. PMID: 12464008 Free PMC article.
-
Manipulation of iron to determine survival: competition between host and pathogen.Immunol Res. 2004;30(1):15-28. doi: 10.1385/IR:30:1:015. Immunol Res. 2004. PMID: 15258308 Review.
References
-
- Bothwell T H, Charlton R W, Motulski A G. In: The Metabolic and Molecular Bases of Inherited Disease. Scriver C R, Beaudet A L, Sly W S, Valle D, editors. New York: McGraw–Hill; 1995. pp. 2237–2269.
-
- Feder J N, Gnirke A, Thomas W, Tsuchihashi Z, Ruddy D A, Basava A, Dormishian F, Domingo R, Ellis M C, Fullan A, et al. Nat Genet. 1996;13:399–408. - PubMed
-
- Cuthbert J A. J Invest Med. 1997;45:518–529. - PubMed
-
- Cox T M, Kelly A L. Curr Opin Genet Dev. 1998;8:274–281. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
