

doi: 10.1101/gad.13.9.1073.
c-Myb is essential for early T cell development
Affiliations
- PMID: 10323859
- PMCID: PMC316940
- DOI: 10.1101/gad.13.9.1073
Item in Clipboard
c-Myb is essential for early T cell development
Genes Dev.
.
Abstract
The c-Myb transcription factor is important for fetal hematopoiesis and has been proposed to mediate later stages of lymphocyte development. Using homozygous null c-Myb/Rag1 chimeric mice, we have determined that c-Myb plays an important role in the differentiation of macrophages and lymphocytes from precursor stem cells. We also determine that deletion of c-Myb leads to a complete block in early T cell development just before the oligopotent thymocyte matures into the definitive T cell precursor. These data indicate that c-Myb plays an important role at multiple stages of hematopoiesis and is required at an early stage of T cell development.
Figures
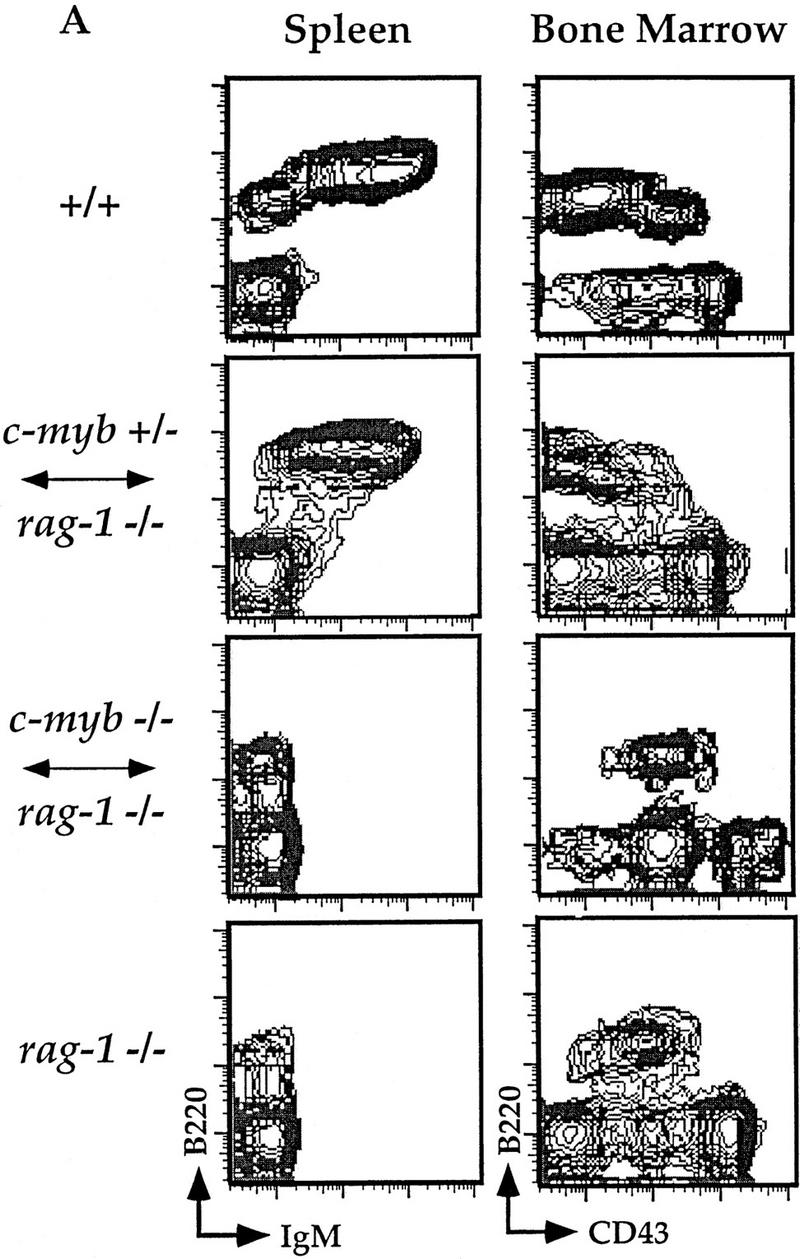
Flow cytometric analyses of B and T lymphocytes from Rag1−/− ↔ c-Myb−/− mice. Splenocytes and thymocytes were isolated from Rag1−/− ↔ c-Myb−/−, Rag1−/− ↔ c-Myb+/−, Rag1−/−, and 129 control mice and identified with combinations of B and T lineage-specific antibodies (see text for details). Chimera 2 was used as an example; similar results were obtained from all other chimeras (data not shown). (A) Splenic B cells and bone marrow B cell precursors; (B) splenic and thymic T cells.
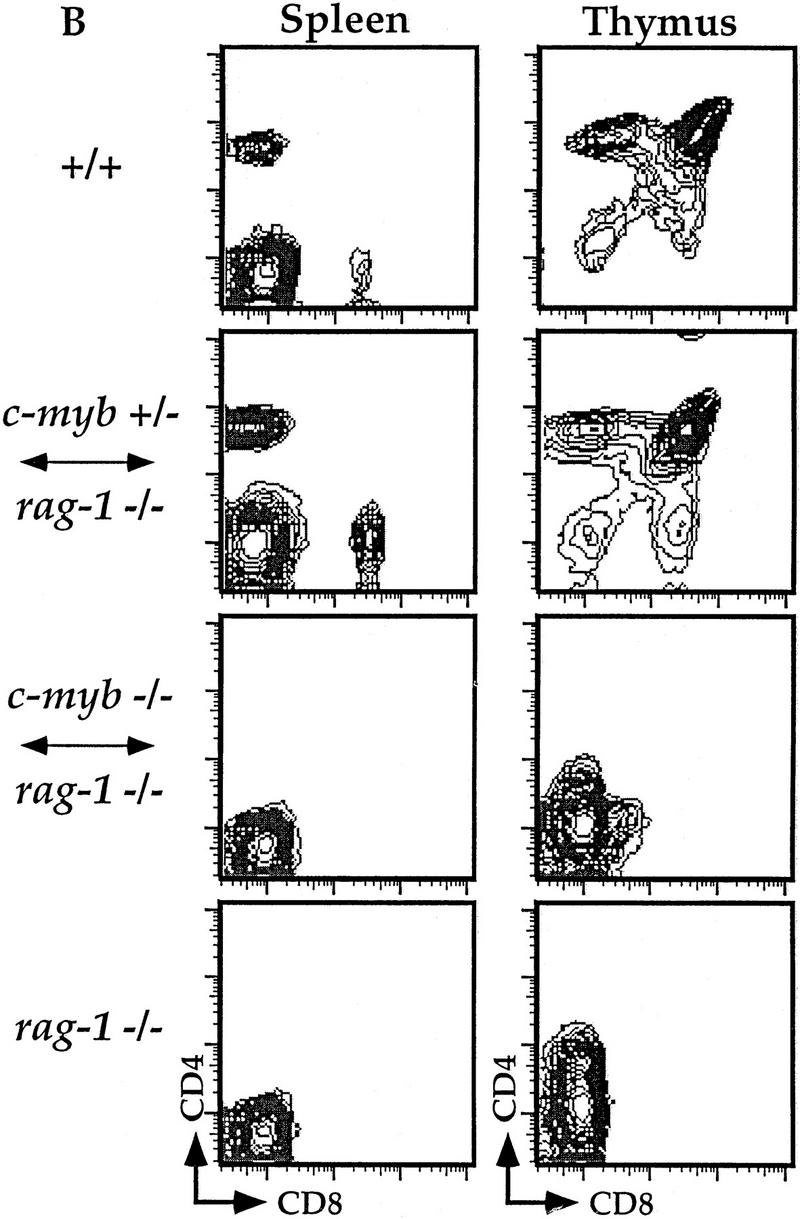
Flow cytometric analyses of B and T lymphocytes from Rag1−/− ↔ c-Myb−/− mice. Splenocytes and thymocytes were isolated from Rag1−/− ↔ c-Myb−/−, Rag1−/− ↔ c-Myb+/−, Rag1−/−, and 129 control mice and identified with combinations of B and T lineage-specific antibodies (see text for details). Chimera 2 was used as an example; similar results were obtained from all other chimeras (data not shown). (A) Splenic B cells and bone marrow B cell precursors; (B) splenic and thymic T cells.
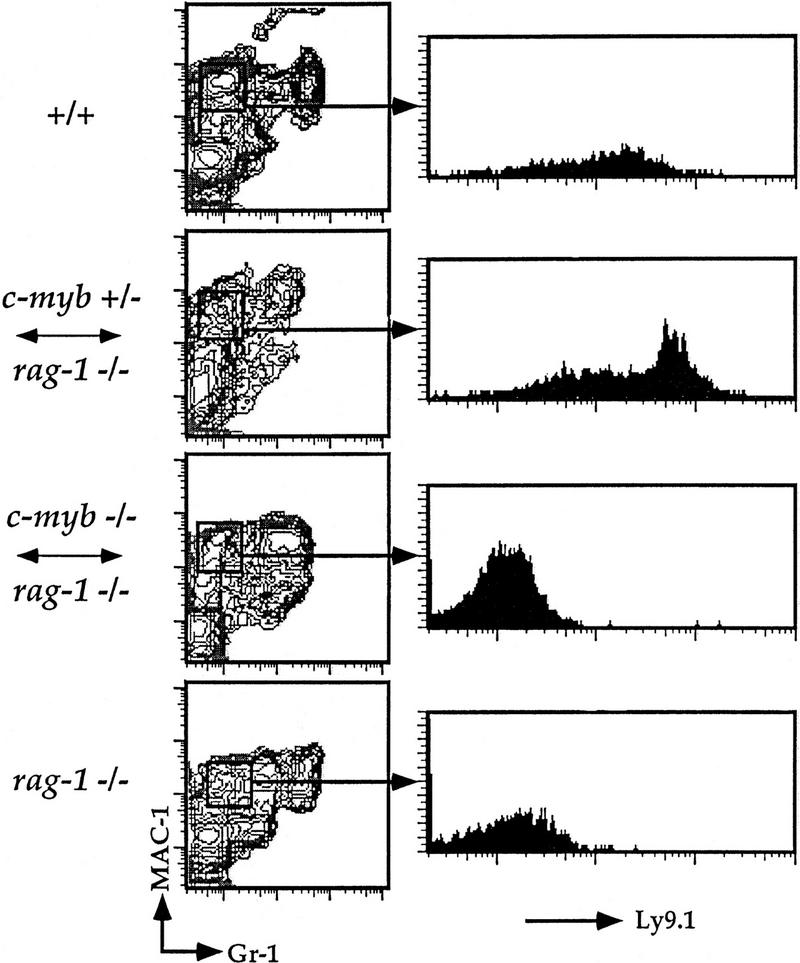
Flow cytometric analysis of splenic macrophages.Splenocytes were isolated from Rag1−/− ↔ c-Myb−/−, Rag1−/− ↔ c-Myb+/−, Rag1−/−, and 129 control mice and identified with antibodies against the macrophage lineage markers Mac-1 and Gr-1 and the c-Myb−/− cell-specific marker Ly9.1 (see text ). The mature macrophage population was gated on and analyzed for Ly9.1 expression as shown. Chimera 2 is shown; similar results were obtained from all other chimeras (data not shown).
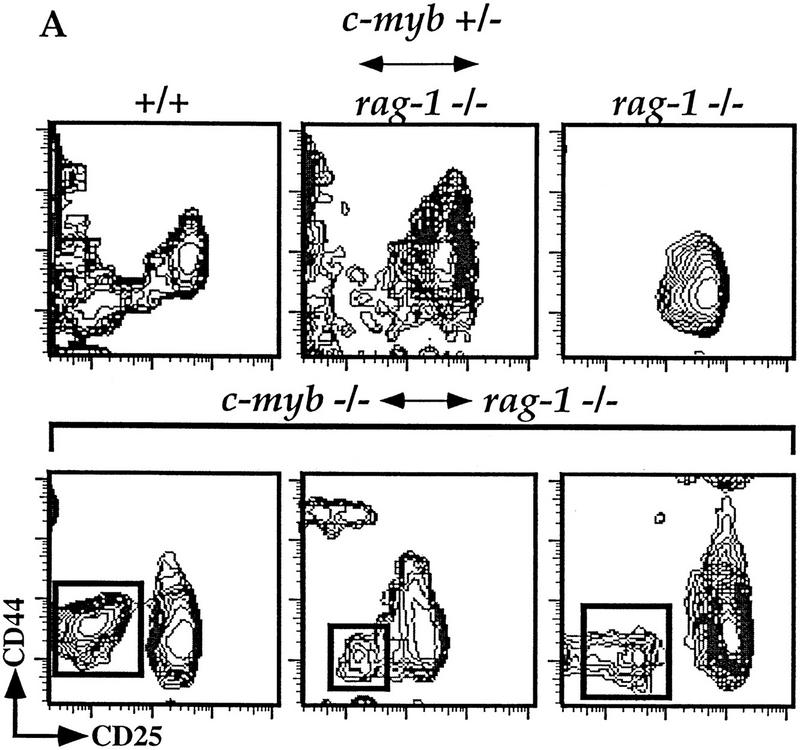
c-Myb−/− HSCs can generate early double-negative thymocytes. (A) Thymocytes were isolated from representative Rag1−/− ↔ c-Myb+/−, Rag1−/−, phenotype positive Rag1−/− ↔ c-Myb−/−, and 129 control mice and analyzed for surface expression of T-lineage-specific markers. Cells expressing any one of the CD8/CD3/B220/Mac-1/Gr-1/Ter-119 markers were excluded from the analysis and the remainder plotted based on their expression of CD25 (x-axis) and CD44 (y-axis). For the Rag1−/− ↔ c-Myb−/− mice, representative phenotype-positive data are shown. (B) Analysis gates drawn on the CD44loCD25− and CD44+/loCD25+ cells for analysis of the expression of the 129 lineage-specific surface marker Ly9.1 are shown.
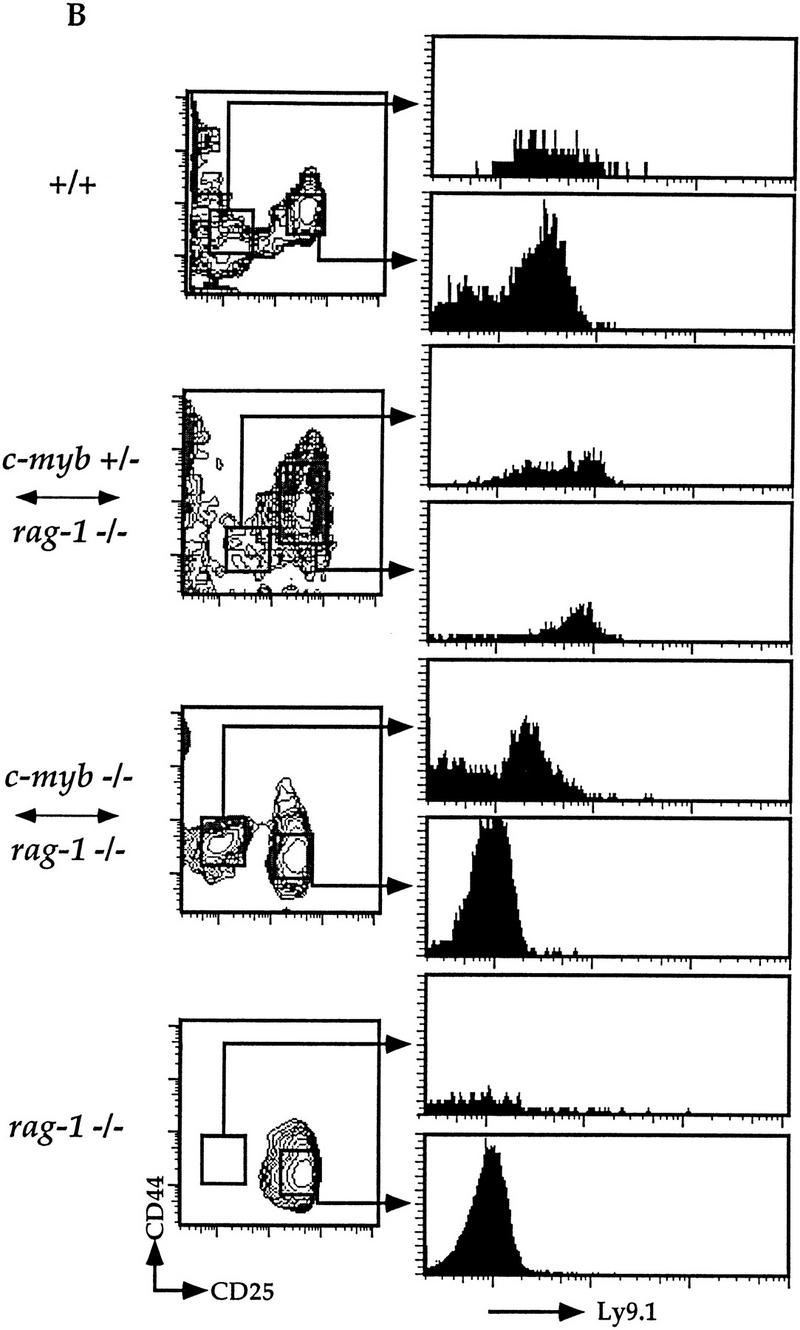
c-Myb−/− HSCs can generate early double-negative thymocytes. (A) Thymocytes were isolated from representative Rag1−/− ↔ c-Myb+/−, Rag1−/−, phenotype positive Rag1−/− ↔ c-Myb−/−, and 129 control mice and analyzed for surface expression of T-lineage-specific markers. Cells expressing any one of the CD8/CD3/B220/Mac-1/Gr-1/Ter-119 markers were excluded from the analysis and the remainder plotted based on their expression of CD25 (x-axis) and CD44 (y-axis). For the Rag1−/− ↔ c-Myb−/− mice, representative phenotype-positive data are shown. (B) Analysis gates drawn on the CD44loCD25− and CD44+/loCD25+ cells for analysis of the expression of the 129 lineage-specific surface marker Ly9.1 are shown.
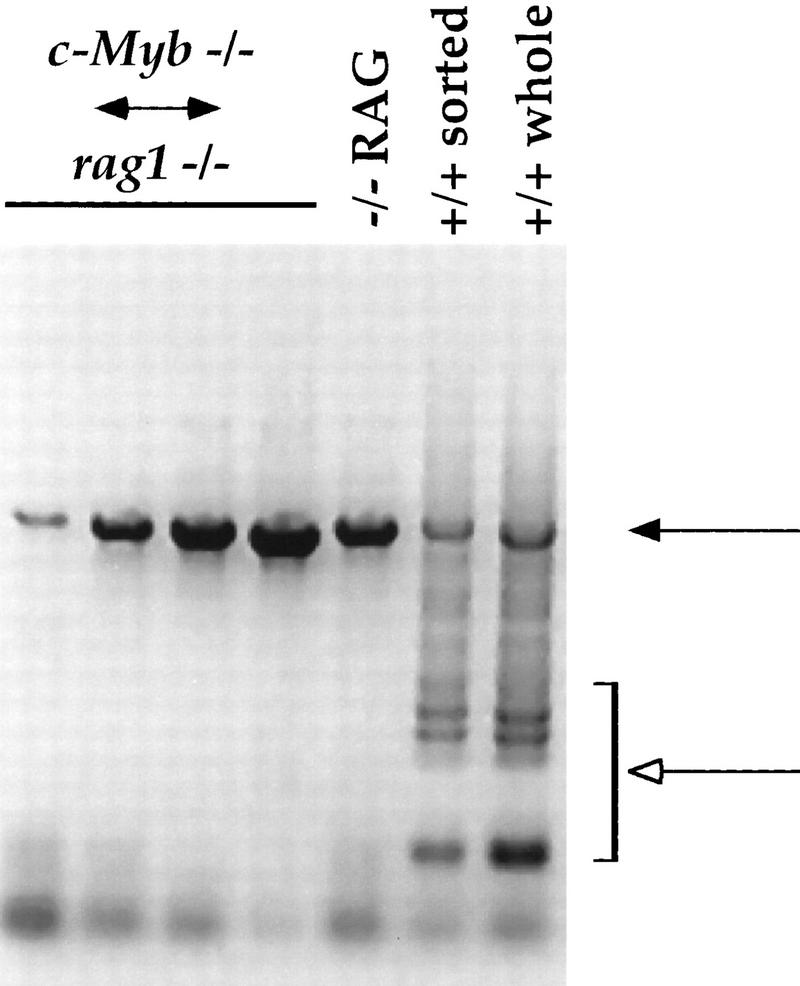
TCRβ-chain gene rearrangement in thymocytes from the Rag1−/− ↔ c-Myb−/− mice. DNA was purified from thymuses from the Rag1−/− ↔ c-Myb−/− and Rag1−/− mice and from both whole thymus and sorted CD4−CD8−CD3− thymocytes from +/+ 129 mice and analyzed them for TCR β-chain gene rearrangement as described previously (Anderson et al., 1992). Results obtained from the Rag1−/− ↔ c-Myb−/− chimeras 1, 2, 9, and 11 (left to right) are shown and are representative of the other chimeras. (Solid arrow) Germ-line band; (open arrow) Dβ–Jβ-rearranged bands.
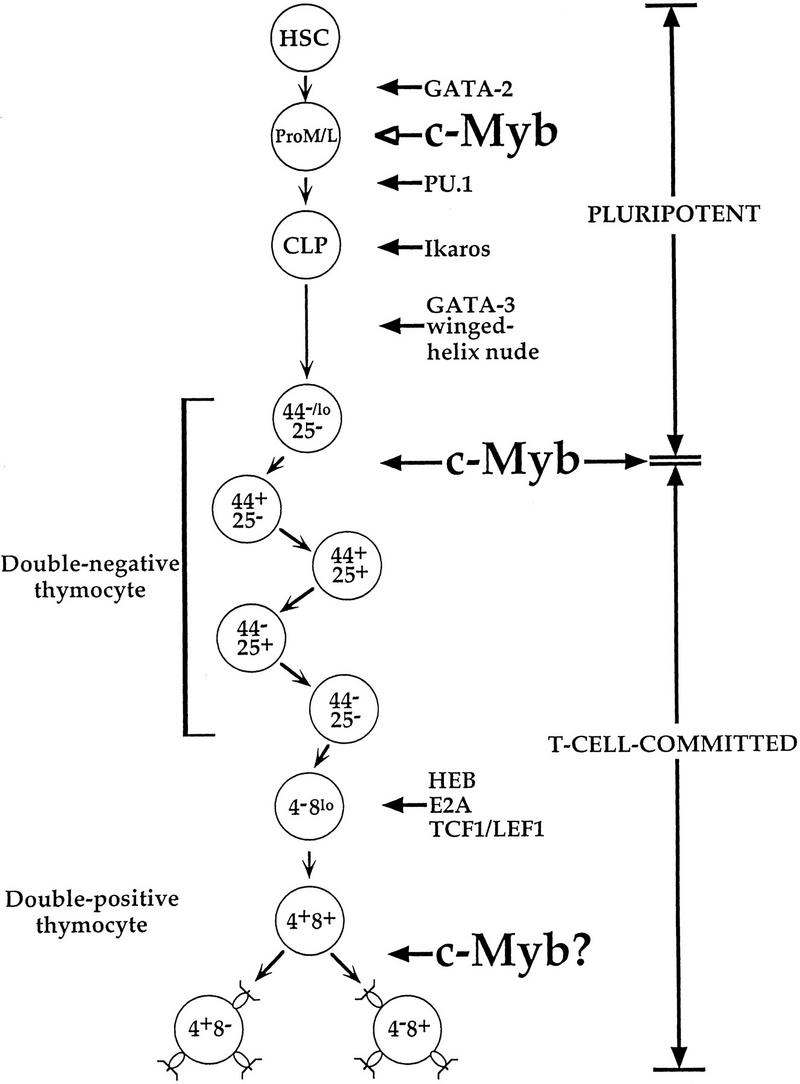
Transcription factors important in early and intermediate T cell development; data are from Glimcher and Singh (1999) and references therein.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases