

UASrpg can function as a heterochromatin boundary element in yeast
- PMID: 10323861
- PMCID: PMC316950
- DOI: 10.1101/gad.13.9.1089
UASrpg can function as a heterochromatin boundary element in yeast
Abstract
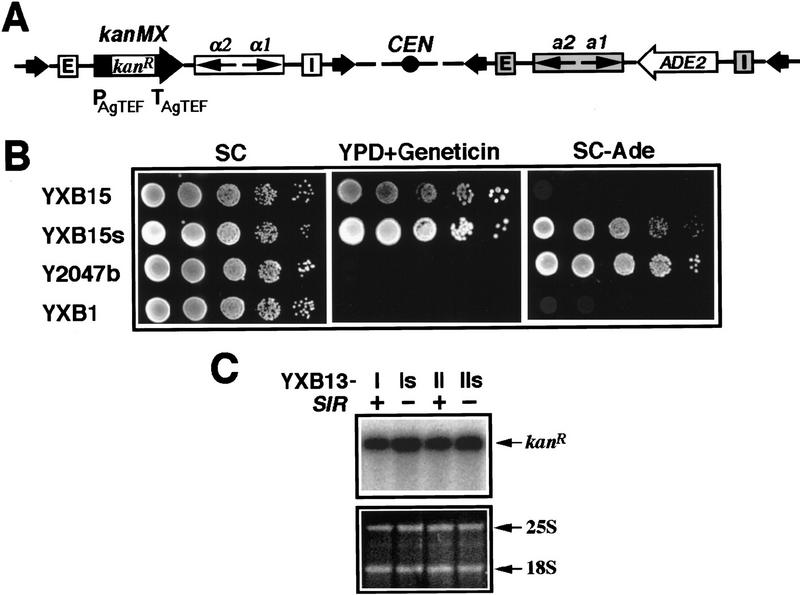
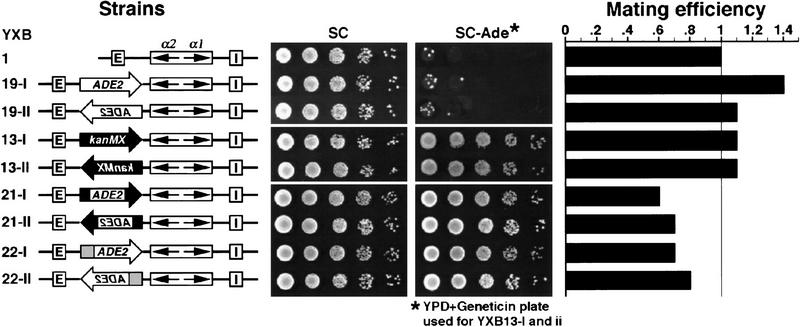
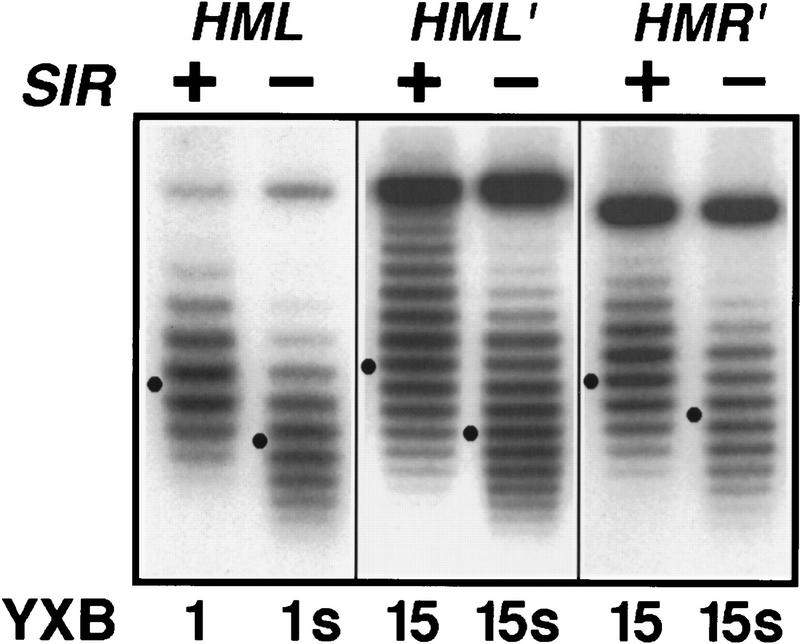
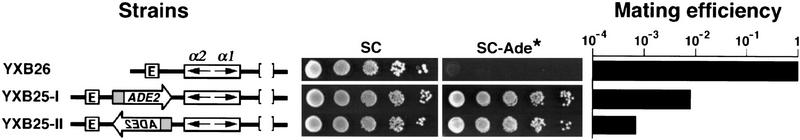
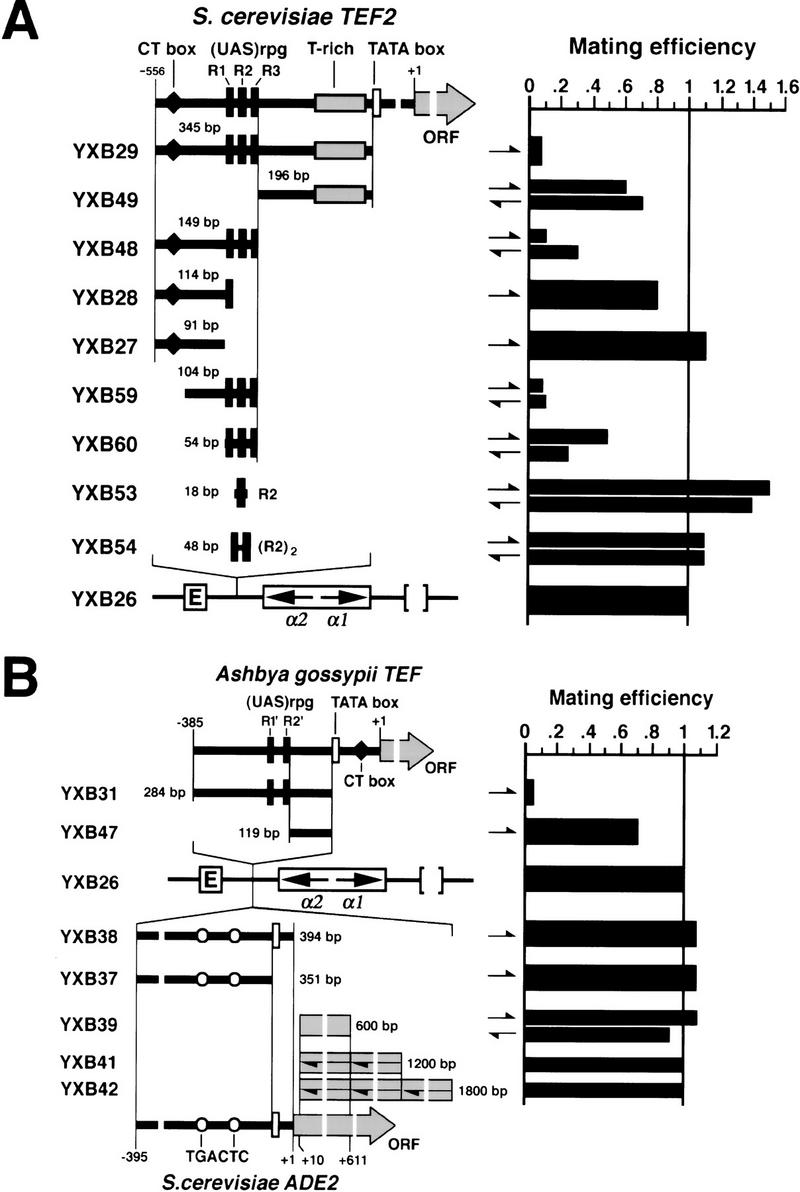
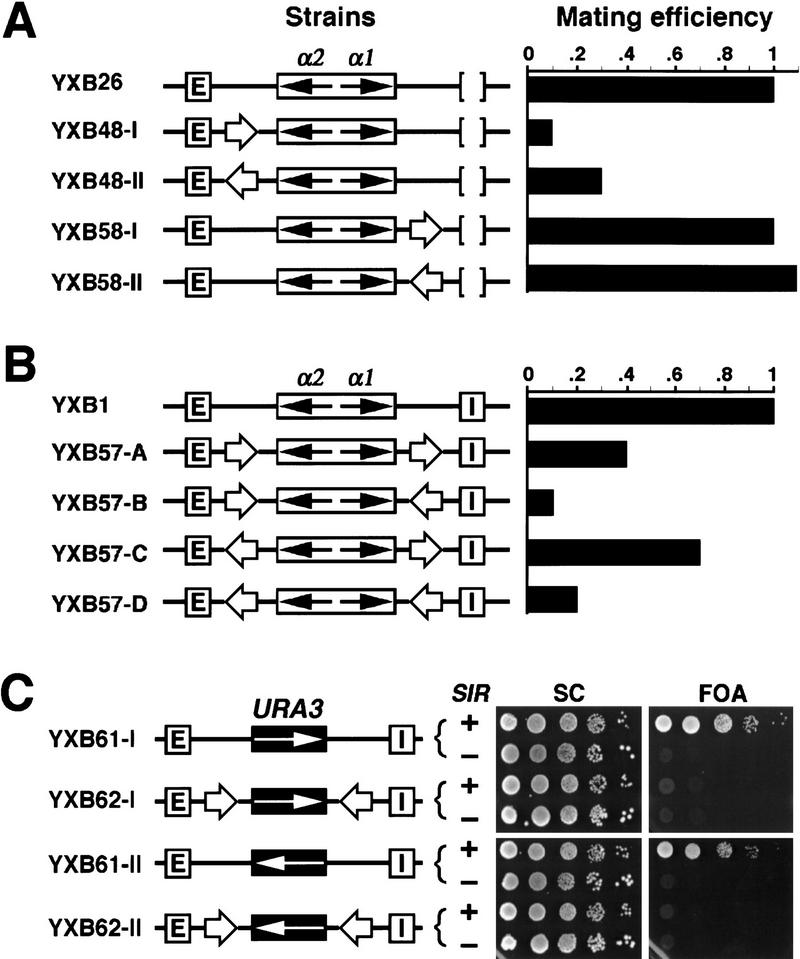
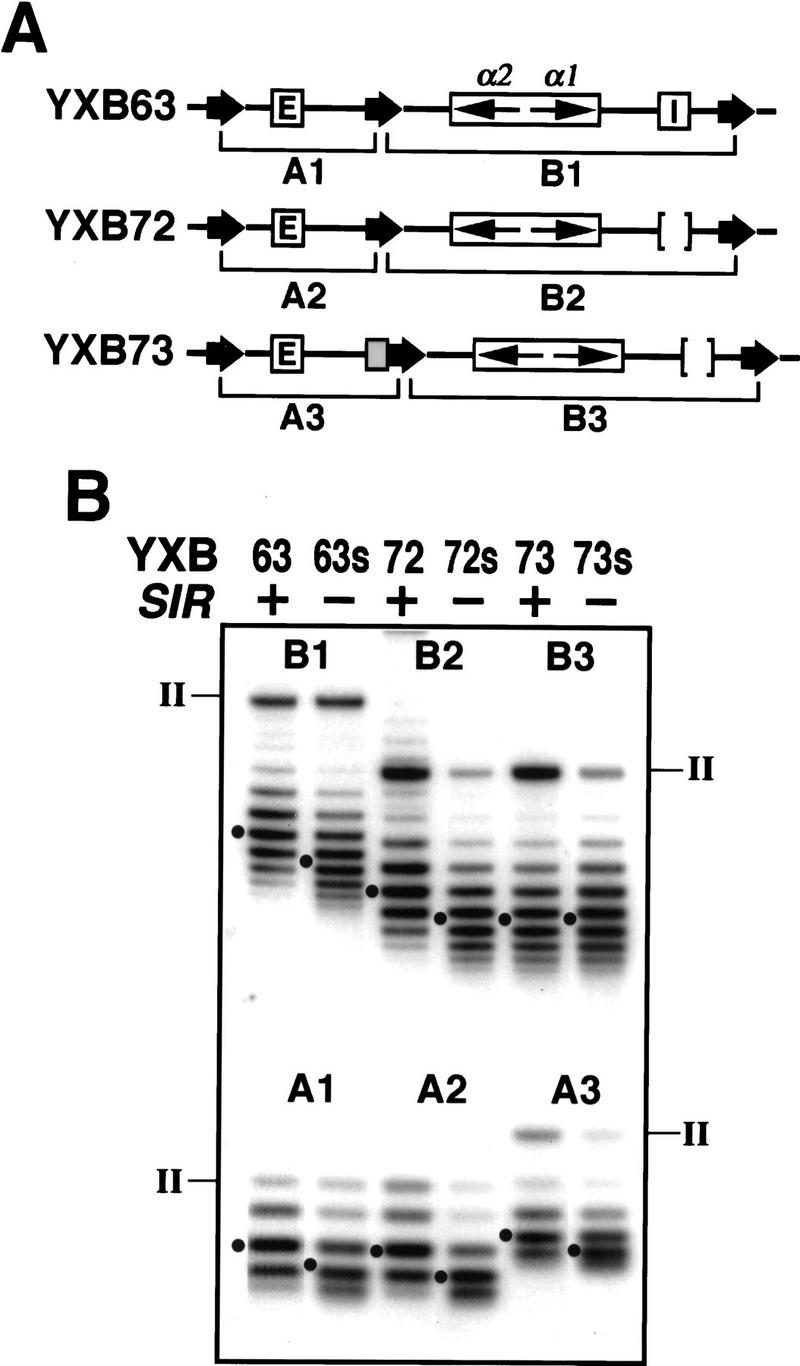
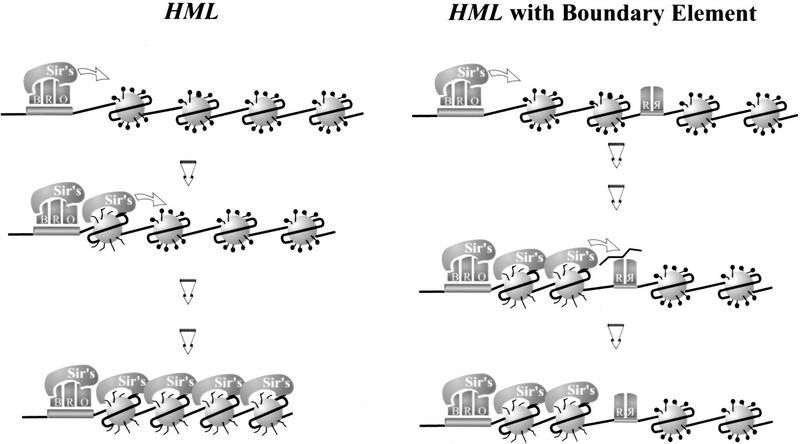
The HM loci in Saccharomyces cerevisiae constitute region-specific but gene-nonspecific repression domains, as a number of heterologous genes transcribed by RNA polymerase II or III are silenced when placed at these loci. The promoters of the Ashbya gossypii TEF gene and the S. cerevisiae TEF1 and TEF2 genes, however, are resistant to transcriptional silencing by the HM silencers in yeast. Moreover, when interposed between the HML alpha genes and the E silencer, certain segments of these promoters block the repression effect of the silencer on the alpha genes. All of these fragments contain UASrpg (upstream activation sequence of ribosome protein genes) composed of multiple binding sites for Rap1. In fact, a 149-bp segment consisting essentially of only three tandem Rap1-binding sites from the UASrpg of yeast TEF2 exhibits silencer-blocking activity. This element also exhibits insulating activity and orientation dependence characteristic of known chromatin boundary elements. Finally, the element blocks the physical spread of heterochromatin initiated at a silencer. This segment provides the first example of chromatin domain boundary or insulator elements in yeast.
Figures
References
-
- Abraham J, Nasmyth KA, Strathern JN, Klar AJS, Hicks JB. Regulation of mating type information in yeast: negative control requiring sequences both 5′ and 3′ to the regulated region. J Mol Biol. 1984;176:307–331. - PubMed
-
- Aparicio OM, Gottschling DE. Overcoming telomeric silencing: A transactivator competes to establish gene expression in a cell cycle-dependent way. Genes & Dev. 1994;8:1133–1146. - PubMed
-
- Brand AH, Breeden L, Abraham J, Sternglanz R, Nasmyth K. Characterization of a ‘silencer’ in yeast: A DNA sequence with properties opposite to those of transcriptional enhancer. Cell. 1985;41:41–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases