

Activation of distinct cAMP-dependent and cGMP-dependent pathways by nitric oxide in cardiac myocytes
- PMID: 10325239
- PMCID: PMC10183997
- DOI: 10.1161/01.res.84.9.1020
Activation of distinct cAMP-dependent and cGMP-dependent pathways by nitric oxide in cardiac myocytes
Abstract
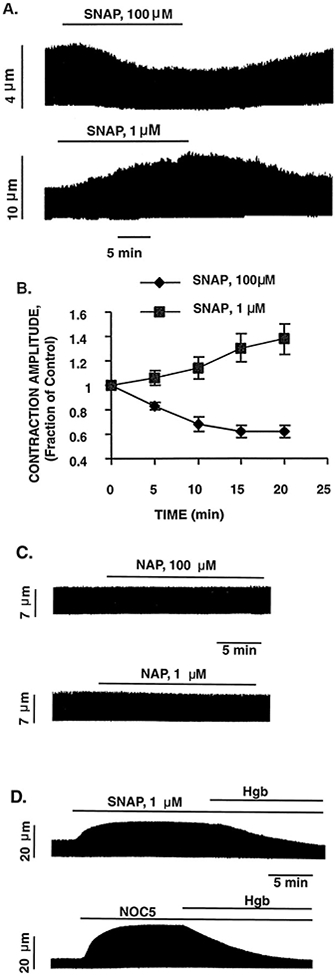
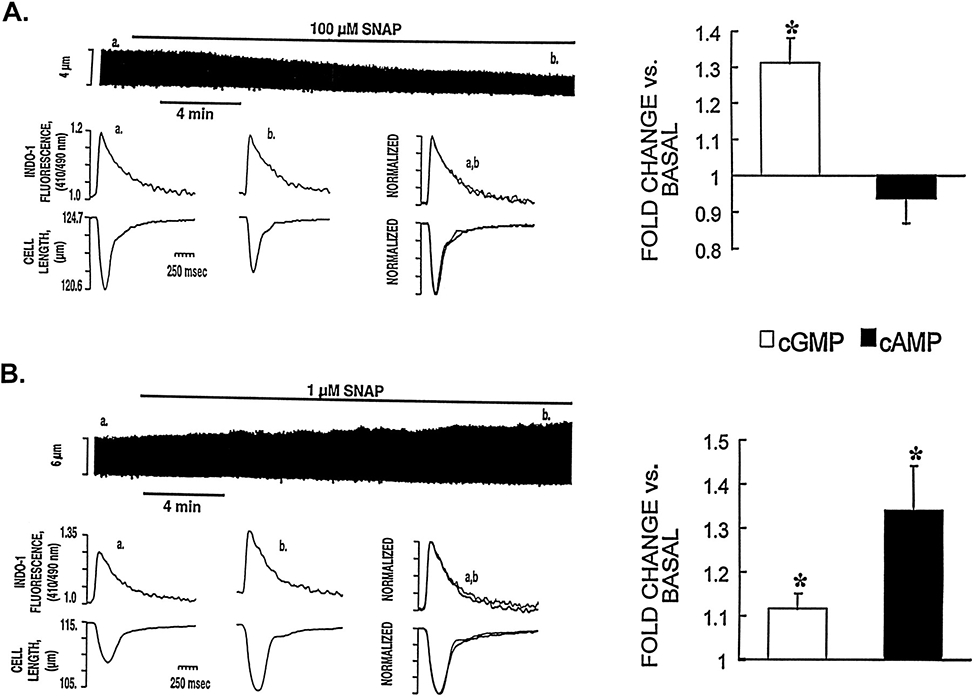
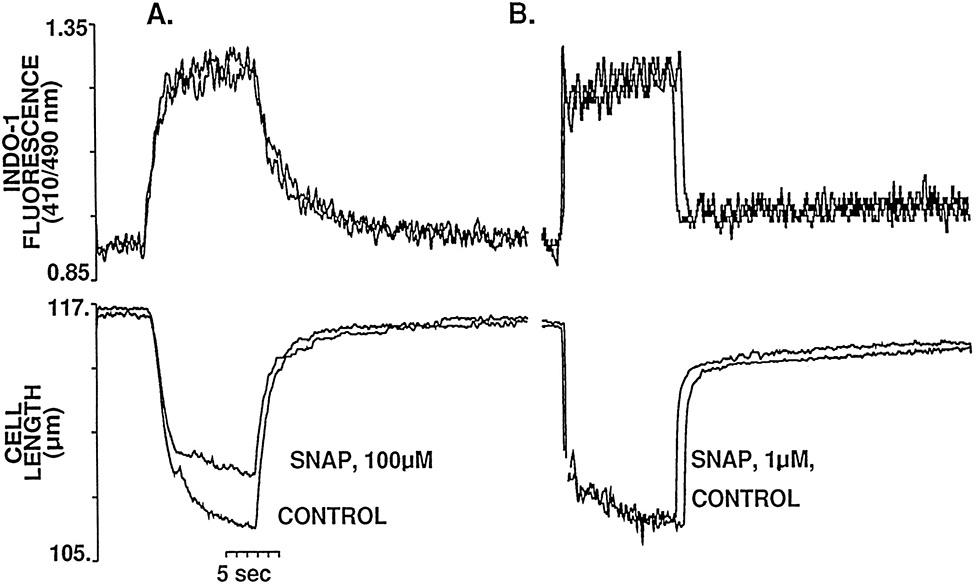
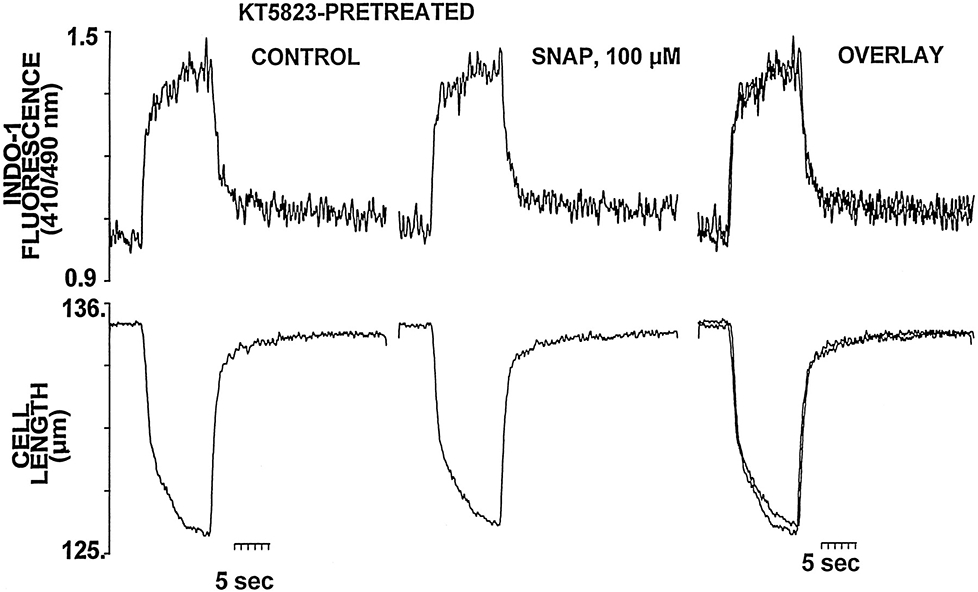
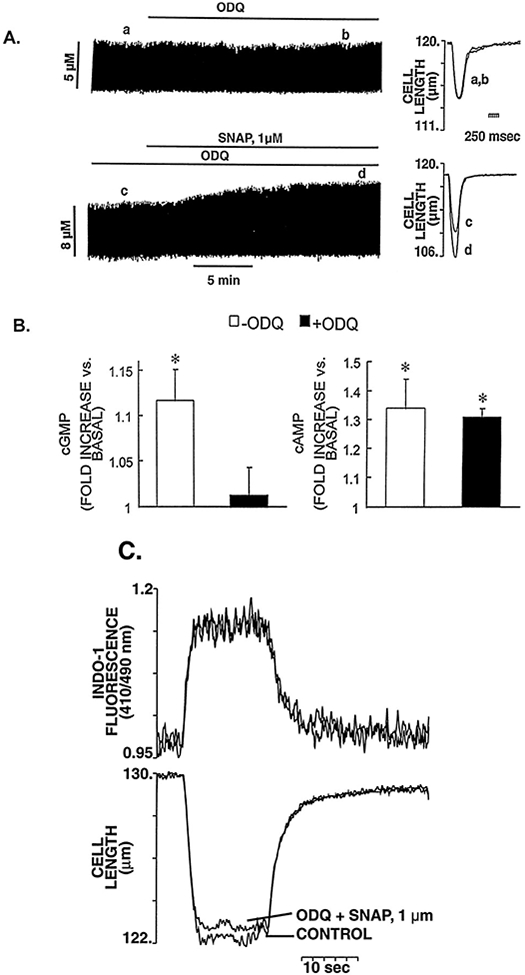
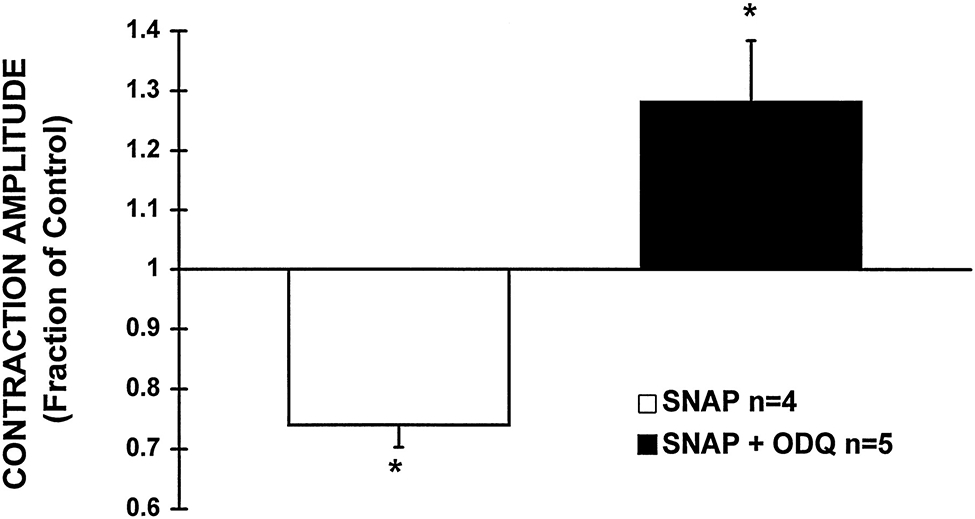
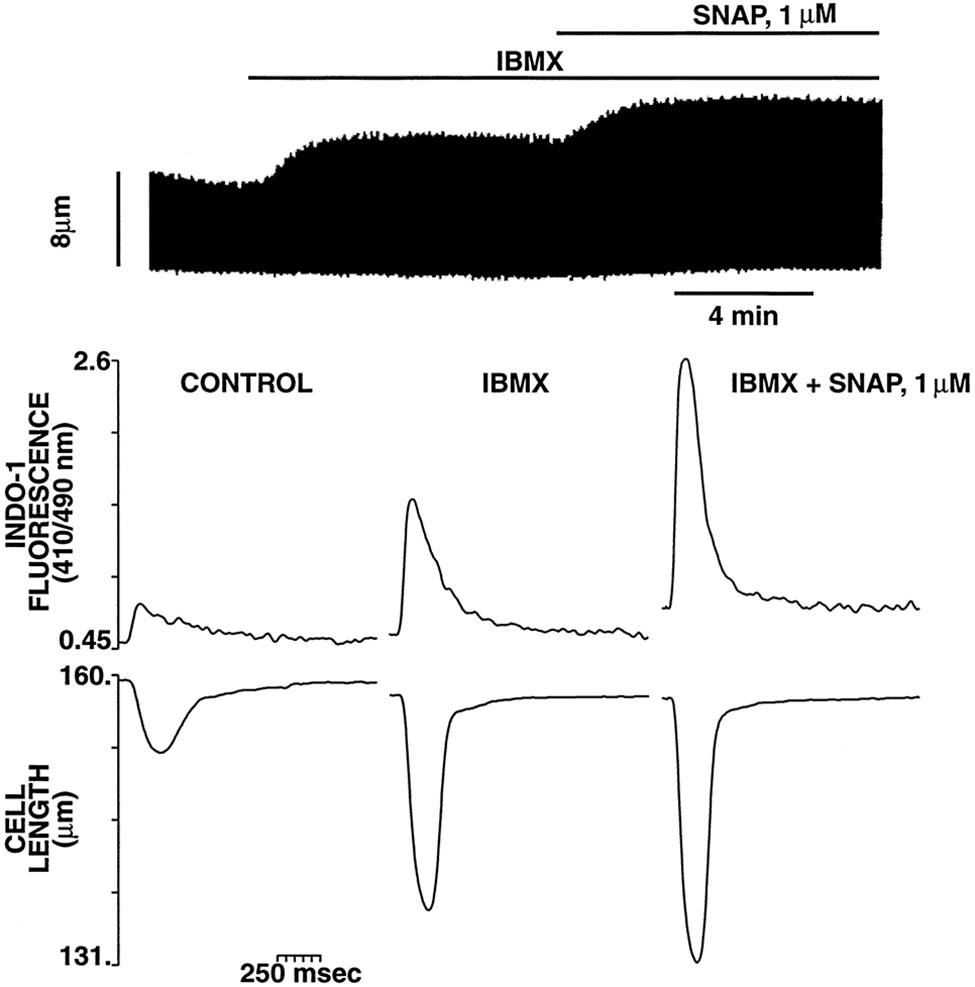
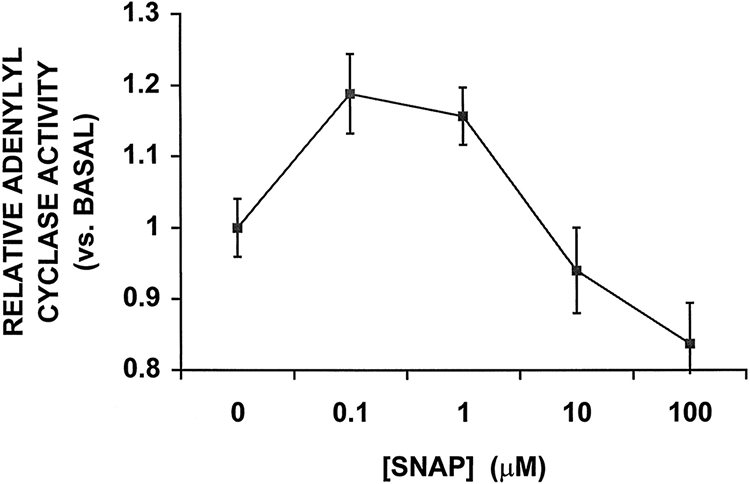
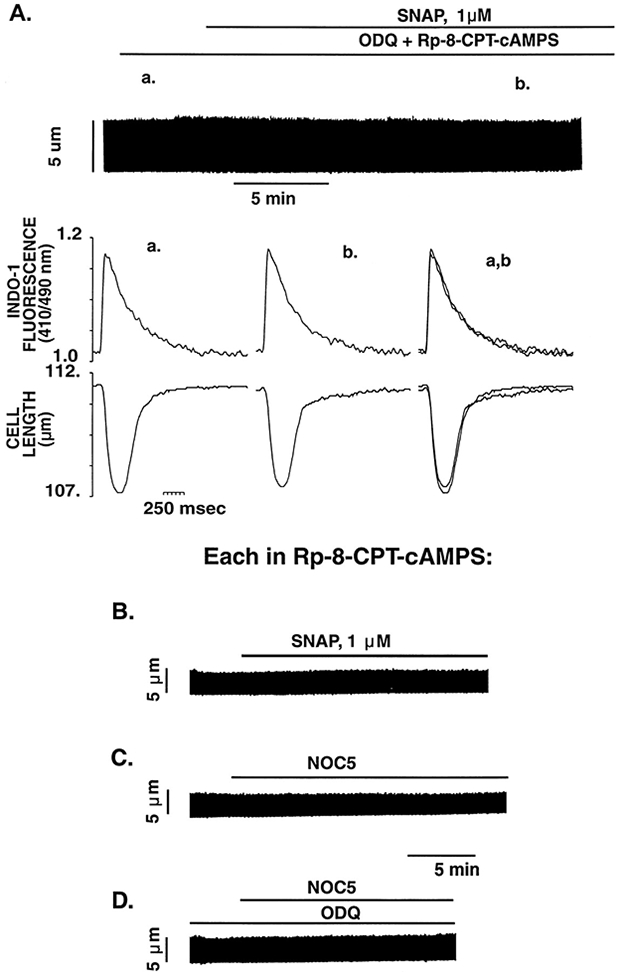
Nitric oxide (NO) donors were recently shown to produce biphasic contractile effects in cardiac tissue, with augmentation at low NO levels and depression at high NO levels. We examined the subcellular mechanisms involved in the opposing effects of NO on cardiac contraction and investigated whether NO modulates contraction exclusively via guanylyl cyclase (GC) activation or whether some contribution occurs via cGMP/PKG-independent mechanisms, in indo 1-loaded adult cardiac myocytes. Whereas a high concentration of the NO donor S-nitroso-N-acetylpenicillamine (SNAP, 100 micromol/L) significantly attenuated contraction amplitude by 24.4+/-4.5% (without changing the Ca2+ transient or total cAMP), a low concentration of SNAP (1 micromol/L) significantly increased contraction amplitude (38+/-10%), Ca2+ transient (26+/-10%), and cAMP levels (from 6.2 to 8.5 pmol/mg of protein). The negative contractile response of 100 micromol/L SNAP was completely abolished in the presence of the specific blocker of PKG KT 5823 (1 micromol/L); the positive contractile response of 1 micromol/L SNAP persisted, despite the presence of the selective inhibitor of GC 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ, 10 micromol/L) alone, but was completely abolished in the presence of ODQ plus the specific inhibitory cAMP analog Rp-8-CPT-cAMPS (100 micromol/L), as well as by the NO scavenger oxyhemoglobin. Parallel experiments in cell suspensions showed significant increases in adenylyl cyclase (AC) activity at low concentrations (0.1 to 1 micromol/L) of SNAP (AC, 18% to 20% above basal activity). We conclude that NO can regulate both AC and GC in cardiac myocytes. High levels of NO induce large increases in cGMP and a negative inotropic effect mediated by a PKG-dependent reduction in myofilament responsiveness to Ca2+. Low levels of NO increase cAMP, at least in part, by a novel cGMP-independent activation of AC and induce a positive contractile response.
Figures
References
-
- Moncada S, Palmer RMJ, Higgs EA. Nitric oxide: physiology, pathology, and pharmacology. Pharmacol Rev. 1991;43:109–142. - PubMed
-
- Ignarro LJ, Kadoitz PJ. The pharmacological role of cyclic GMP in vascular smooth muscle relaxation. Annu Rev Pharmacol Toxicol. 1985;25:171–191. - PubMed
-
- Brady AJB, Warren JB, Poole-Wilson PA, Williams TJ, Harding SE. Nitric oxide attenuates cardiac myocyte contraction. Am J Physiol. 1993;265:H176–H182. - PubMed
-
- Meulemans AL, Sipido KR, Sys SU, Brutsaert DL. Atriopeptin III induces early relaxation of ventricular cardiac muscle. Circ Res. 1988;62:1171–1174. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
