

Differential and inefficient splicing of a broadly expressed Drosophila erect wing transcript results in tissue-specific enrichment of the vital EWG protein isoform
- PMID: 10330140
- PMCID: PMC104359
- DOI: 10.1128/MCB.19.6.3998
Differential and inefficient splicing of a broadly expressed Drosophila erect wing transcript results in tissue-specific enrichment of the vital EWG protein isoform
Abstract
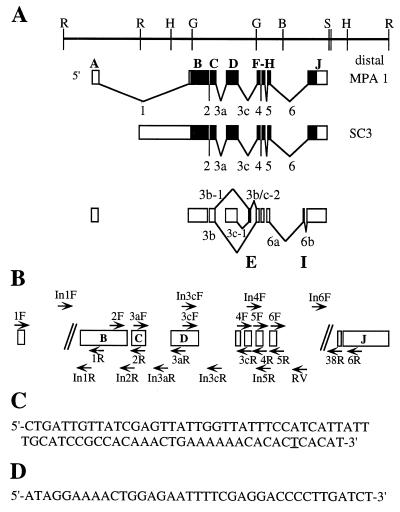
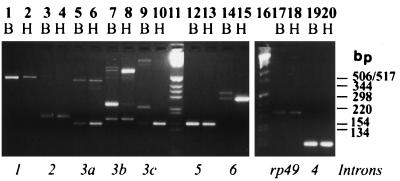
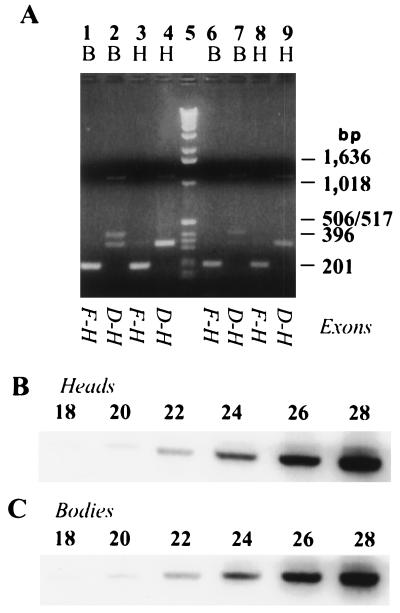
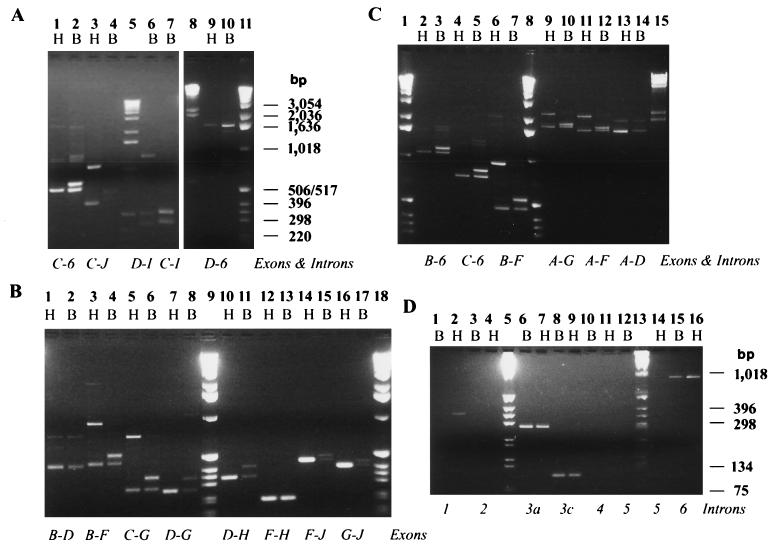
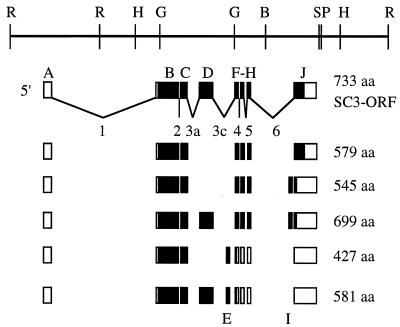
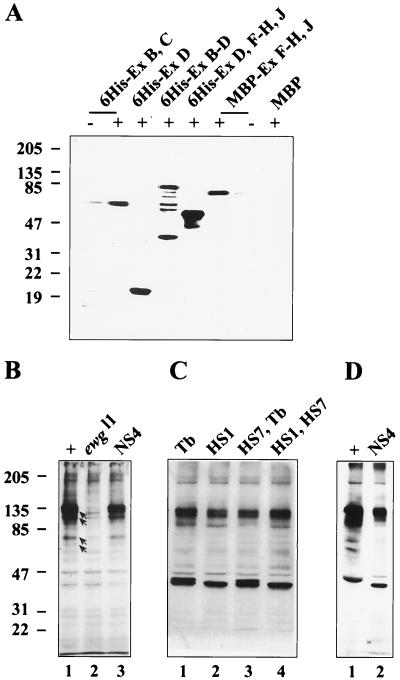
In this report, we document an unusual mode of tissue-enriched gene expression that is primarily mediated by alternative and inefficient splicing. We have analyzed posttranscriptional regulation of the Drosophila erect wing gene, which provides a vital neuronal function and is essential for the formation of certain muscles. Its predominant protein product, the 116-kDa EWG protein, a putative transcriptional regulator, can provide all known erect wing-associated functions. Moreover, consistent with its function, the 116-kDa protein is highly enriched in neurons and is also observed transiently in migrating myoblasts. In contrast to the protein distribution, we observed that erect wing transcripts are present in comparable levels in neuron-enriched heads and neuron-poor bodies of adult Drosophila. Our analyses shows that erect wing transcript consists of 10 exons and is alternatively spliced and that a subset of introns are inefficiently spliced. We also show that the 116-kDa EWG protein-encoding splice isoform is head enriched. In contrast, bodies have lower levels of transcripts that can encode the 116-kDa protein and greater amounts of unprocessed erect wing RNA. Thus, the enrichment of the 116-kDa protein in heads is ensured by tissue-specific alternative and inefficient splicing and not by transcriptional regulation. Furthermore, this regulation is biologically important, as an increased level of the 116-kDa protein outside the nervous system is lethal.
Figures
Similar articles
-
The neuron-enriched splicing pattern of Drosophila erect wing is dependent on the presence of ELAV protein.Mol Cell Biol. 2000 Mar;20(5):1836-45. doi: 10.1128/MCB.20.5.1836-1845.2000. Mol Cell Biol. 2000. PMID: 10669758 Free PMC article.
-
ERECT WING, the Drosophila member of a family of DNA binding proteins is required in imaginal myoblasts for flight muscle development.Development. 1996 Jan;122(1):31-9. doi: 10.1242/dev.122.1.31. Development. 1996. PMID: 8565844
-
The Drosophila erect wing gene, which is important for both neuronal and muscle development, encodes a protein which is similar to the sea urchin P3A2 DNA binding protein.Mol Cell Biol. 1993 Jun;13(6):3641-9. doi: 10.1128/mcb.13.6.3641-3649.1993. Mol Cell Biol. 1993. PMID: 8388540 Free PMC article.
-
RNA-protein interactions that regulate pre-mRNA splicing.Gene Expr. 2002;10(1-2):79-92. Gene Expr. 2002. PMID: 11868989 Free PMC article. Review.
-
Regulated functional alternative splicing in Drosophila.Nucleic Acids Res. 2012 Jan;40(1):1-10. doi: 10.1093/nar/gkr648. Epub 2011 Sep 8. Nucleic Acids Res. 2012. PMID: 21908400 Free PMC article. Review.
Cited by
-
Memory consolidation in honey bees is enhanced by down-regulation of Down syndrome cell adhesion molecule and changes its alternative splicing.Front Mol Neurosci. 2024 Jan 9;16:1322808. doi: 10.3389/fnmol.2023.1322808. eCollection 2023. Front Mol Neurosci. 2024. PMID: 38264345 Free PMC article.
-
Nutrition controls mitochondrial biogenesis in the Drosophila adipose tissue through Delg and cyclin D/Cdk4.PLoS One. 2009 Sep 9;4(9):e6935. doi: 10.1371/journal.pone.0006935. PLoS One. 2009. PMID: 19742324 Free PMC article.
-
Erect wing regulates synaptic growth in Drosophila by integration of multiple signaling pathways.Genome Biol. 2008 Apr 17;9(4):R73. doi: 10.1186/gb-2008-9-4-r73. Genome Biol. 2008. PMID: 18419806 Free PMC article.
-
Sandhya Koushika: Building new models and communities. Interview with Caitlin Sedwick.J Cell Biol. 2013 Apr 1;201(1):4-5. doi: 10.1083/jcb.2011pi. J Cell Biol. 2013. PMID: 23547027 Free PMC article.
-
The neuron-enriched splicing pattern of Drosophila erect wing is dependent on the presence of ELAV protein.Mol Cell Biol. 2000 Mar;20(5):1836-45. doi: 10.1128/MCB.20.5.1836-1845.2000. Mol Cell Biol. 2000. PMID: 10669758 Free PMC article.
References
-
- Adams M D, Tarng R S, Rio D C. The alternative splicing factor PSI regulates P-element third intron splicing in vivo. Genes Dev. 1997;11:129–138. - PubMed
-
- Becker T S, Burgess S M, Amsterdam A H, Allende M L, Hopkins N. Not really finished is crucial for development of the zebrafish outer retina and encodes a transcription factor highly homologous to human nuclear respiratory factor-1 and avian initiation binding repressor. Development. 1998;125:4369–4378. - PubMed
-
- Calzone F J, Hoog C, Teplow D B, Cutting A E, Zeller R W, Britten R J, Davidson E H. Gene regulatory factors of the sea urchin embryo. I. Purification by affinity chromatography and cloning of P3A2, a novel DNA-binding protein. Development. 1991;112:335–350. - PubMed
-
- Chabot B. Directing alternative splicing: cast and scenarios. Trends Genet. 1996;12:472–478. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases