

Basis for the checkpoint signal specificity that regulates Chk1 and Cds1 protein kinases
- PMID: 10330167
- PMCID: PMC104386
- DOI: 10.1128/MCB.19.6.4262
Basis for the checkpoint signal specificity that regulates Chk1 and Cds1 protein kinases
Abstract
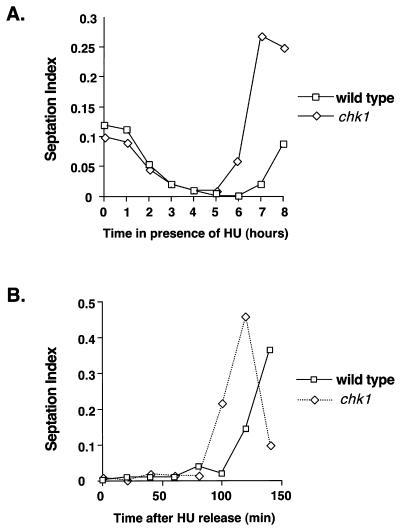
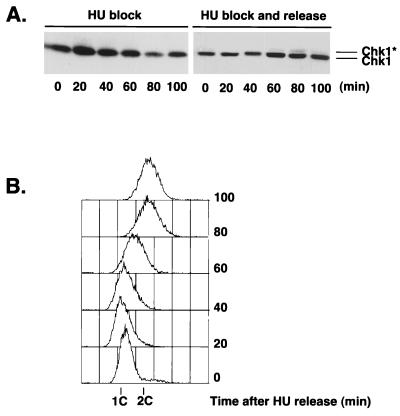
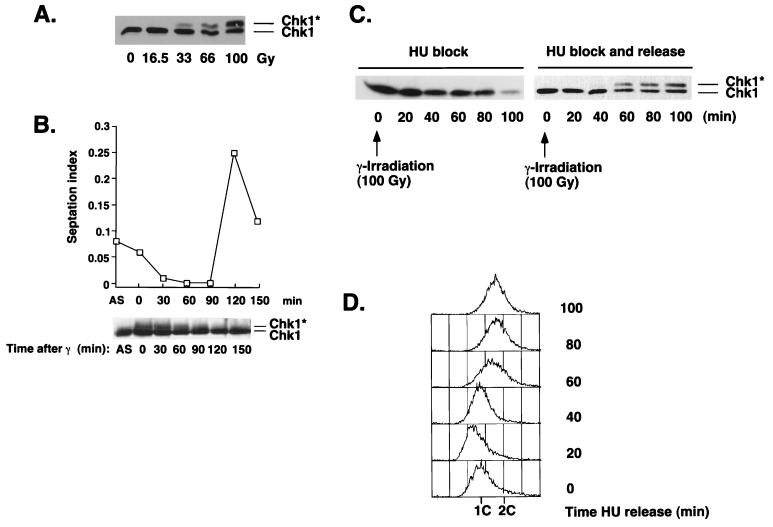
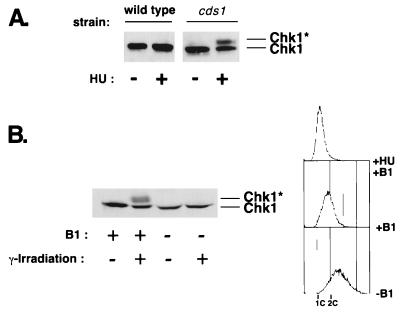
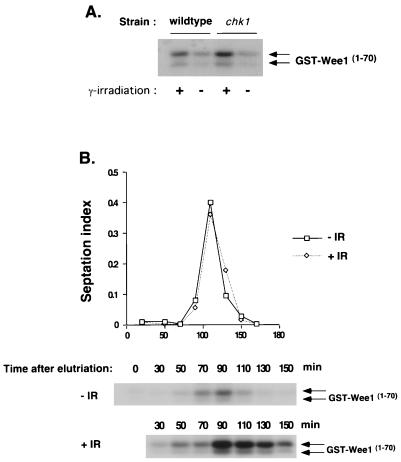
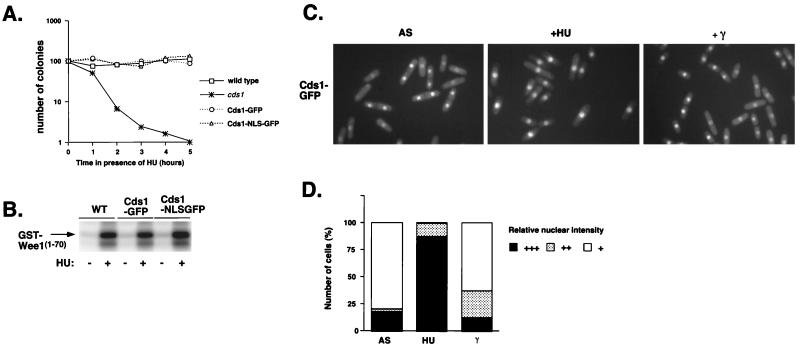
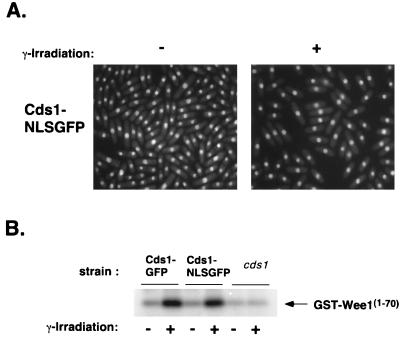
Six checkpoint Rad proteins (Rad1, Rad3, Rad9, Rad17, Rad26, and Hus1) are needed to regulate checkpoint protein kinases Chk1 and Cds1 in fission yeast. Chk1 is required to prevent mitosis when DNA is damaged by ionizing radiation (IR), whereas either kinase is sufficient to prevent mitosis when DNA replication is inhibited by hydroxyurea (HU). Checkpoint Rad proteins are required for IR-induced phosphorylation of Chk1 and HU-induced activation of Cds1. IR activates Cds1 only during the DNA synthesis (S) phase, whereas HU induces Chk1 phosphorylation only in cds1 mutants. Here, we investigate the basis of the checkpoint signal specificity of Chk1 phosphorylation and Cds1 activation. We show that IR fails to induce Chk1 phosphorylation in HU-arrested cells. Release from the HU arrest following IR causes substantial Chk1 phosphorylation. These and other data indicate that Cds1 prevents Chk1 phosphorylation in HU-arrested cells, which suggests that Cds1 actively suppresses a repair process that leads to Chk1 phosphorylation. Cds1 becomes more highly concentrated in the nucleus only during the S phase of the cell cycle. This finding correlates with S-phase specificity of IR-induced activation of Cds1. However, constitutive nuclear localization of Cds1 does not enhance IR-induced activation of Cds1. This result suggests that Cds1 activation requires DNA structures or protein activities that are present only during S phase. These findings help to explain how Chk1 and Cds1 respond to different checkpoint signals.
Figures
References
-
- Aligue R, Wu L, Russell P. Regulation of Schizosaccharomyces pombe Wee1 tyrosine kinase. J Biol Chem. 1997;272:13320–13325. - PubMed
-
- Basi G, Schmid E, Maundrell K. TATA box mutations in the Schizosaccharomyces pombe nmt1 promoter affect transcription efficiency but not the transcription start point or thiamine repressibility. Gene. 1993;123:131–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous