

timrit Lengthens circadian period in a temperature-dependent manner through suppression of PERIOD protein cycling and nuclear localization
- PMID: 10330175
- PMCID: PMC104394
- DOI: 10.1128/MCB.19.6.4343
timrit Lengthens circadian period in a temperature-dependent manner through suppression of PERIOD protein cycling and nuclear localization
Abstract
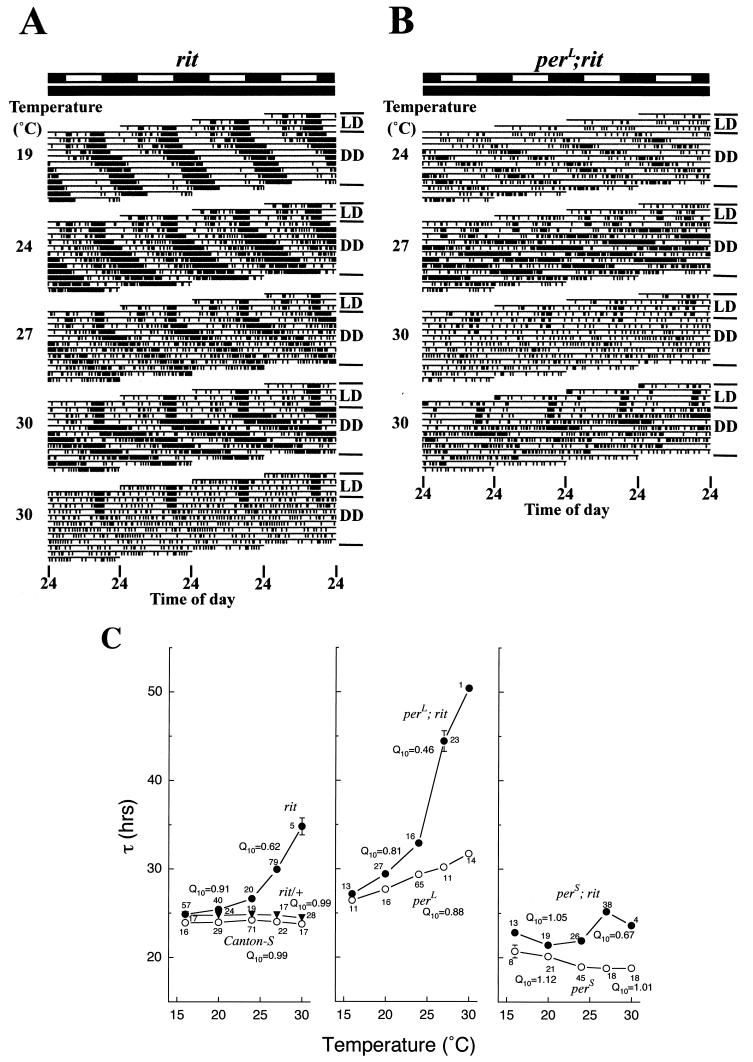
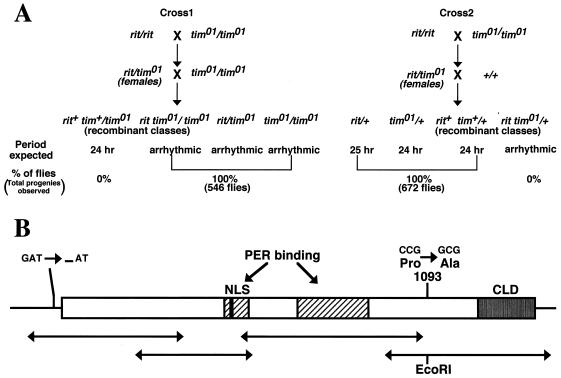
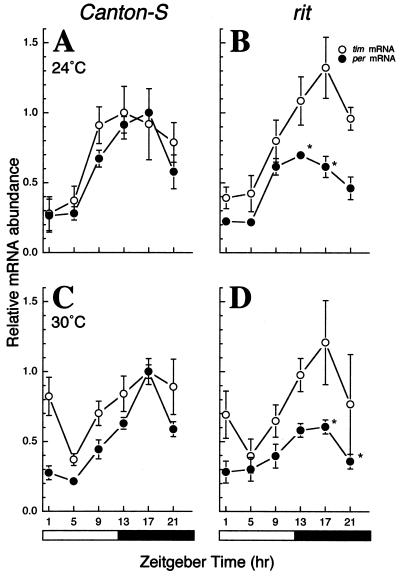
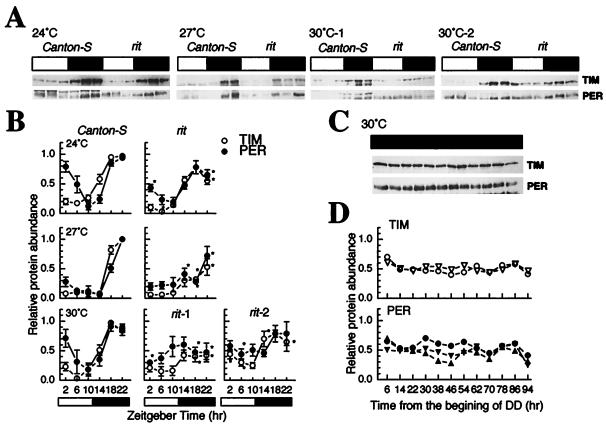
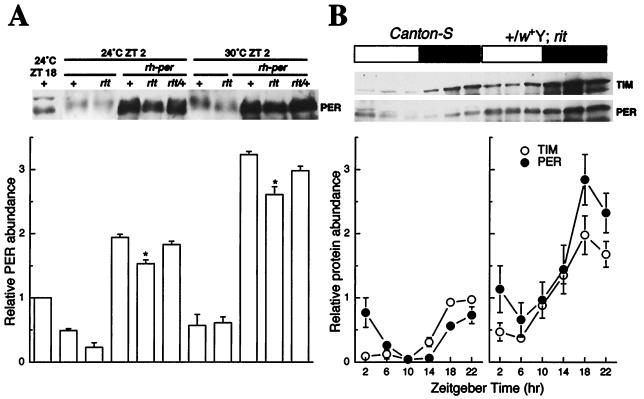
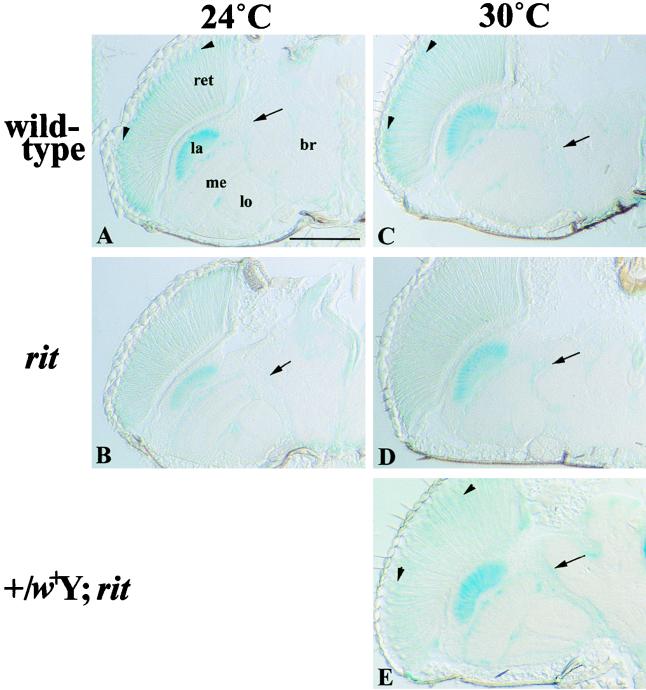
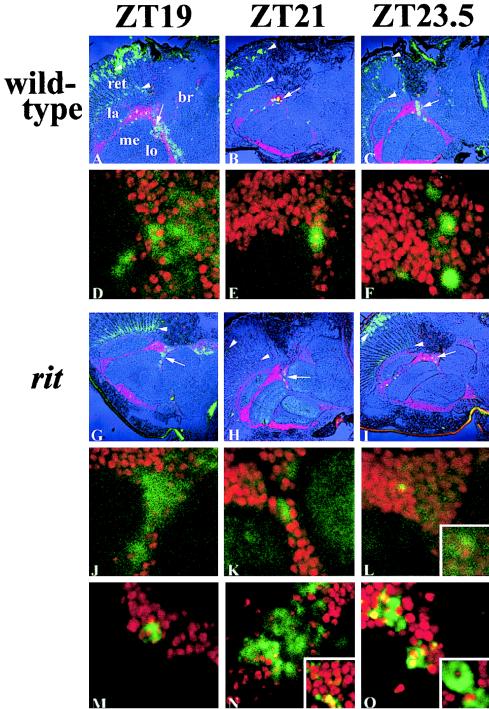
A fundamental feature of circadian clocks is temperature compensation of period. The free-running period of ritsu (timrit) (a novel allele of timeless [tim]) mutants is drastically lengthened in a temperature-dependent manner. PER and TIM protein levels become lower in timrit mutants as temperature becomes higher. This mutation reduces per mRNA but not tim mRNA abundance. PER constitutively driven by the rhodopsin1 promoter is lowered in rit mutants, indicating that timrit mainly affects the per feedback loop at a posttranscriptional level. An excess of per+ gene dosage can ameliorate all rit phenotypes, including the weak nuclear localization of PER, suggesting that timrit affects circadian rhythms by reducing PER abundance and its subsequent transportation into nuclei as temperature increases.
Figures

References
-
- Allada R, White N E, So W V, Hall J C, Rosbash M. A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell. 1998;93:791–804. - PubMed
-
- Baylies M K, Vosshall L B, Sehgal A, Young M W. New short period mutations of the Drosophila clock gene per. Neuron. 1992;9:575–581. - PubMed
-
- Cote G G, Brody S. Circadian rhythms in Drosophila melanogaster: analysis of period as a function of gene dosage at the per (period) locus. J Theor Biol. 1986;121:487–503. - PubMed
-
- Curtin K D, Huang Z J, Rosbash M. Temporally regulated nuclear entry of the Drosophila period protein contributes to the circadian clock. Neuron. 1995;14:365–372. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases