

Cooperative interaction between GATA-4 and GATA-6 regulates myocardial gene expression
- PMID: 10330176
- PMCID: PMC104395
- DOI: 10.1128/MCB.19.6.4355
Cooperative interaction between GATA-4 and GATA-6 regulates myocardial gene expression
Abstract
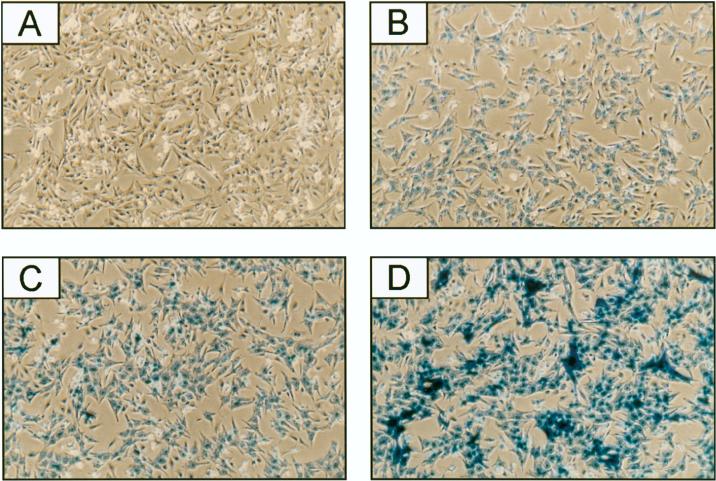
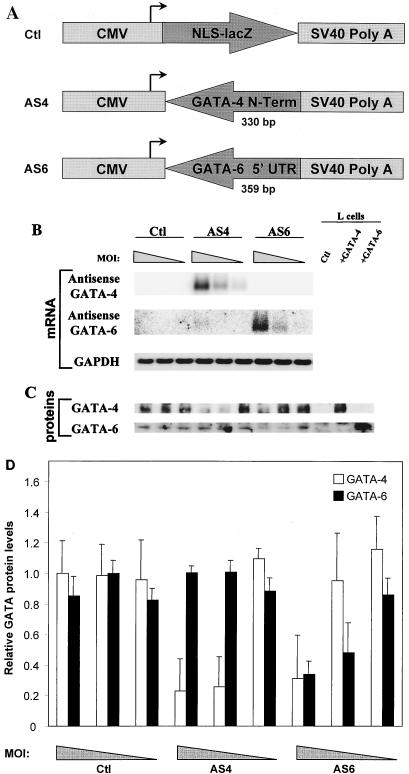
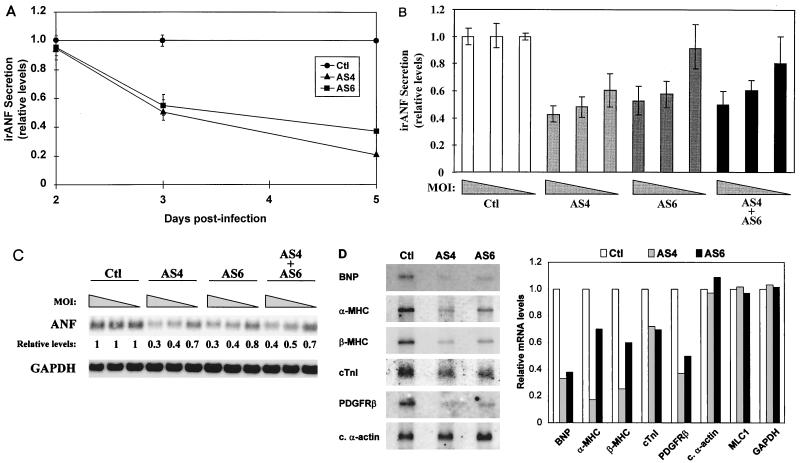
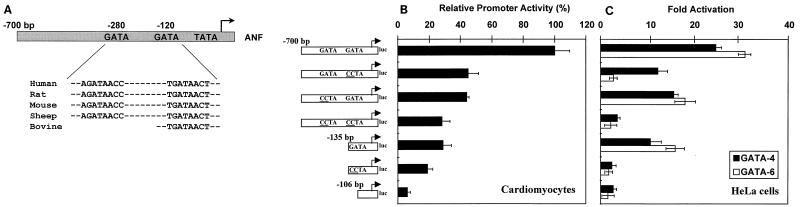
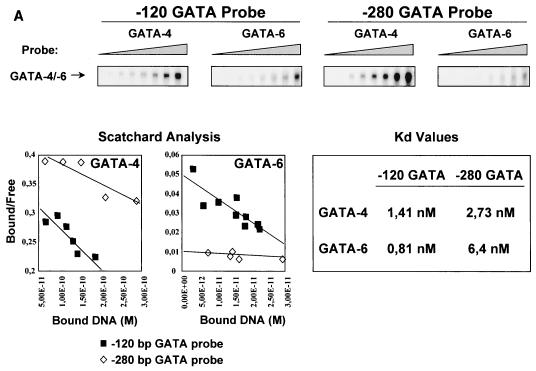
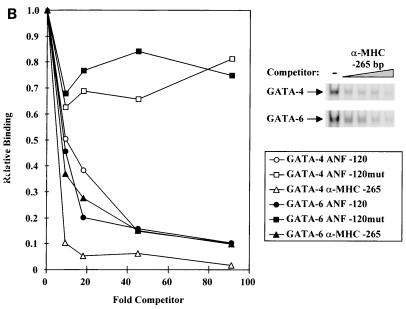
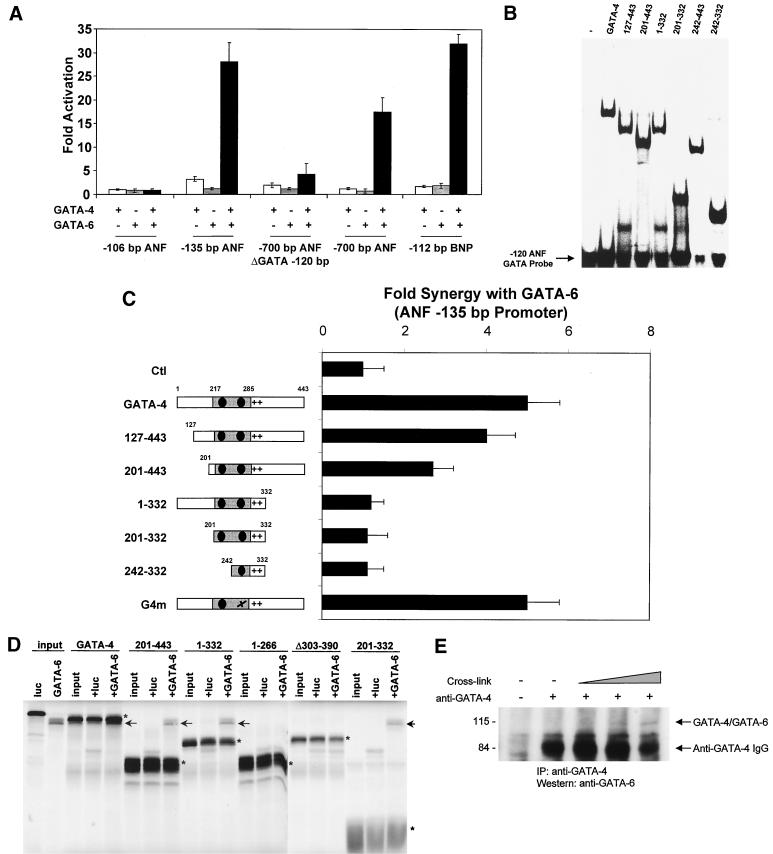
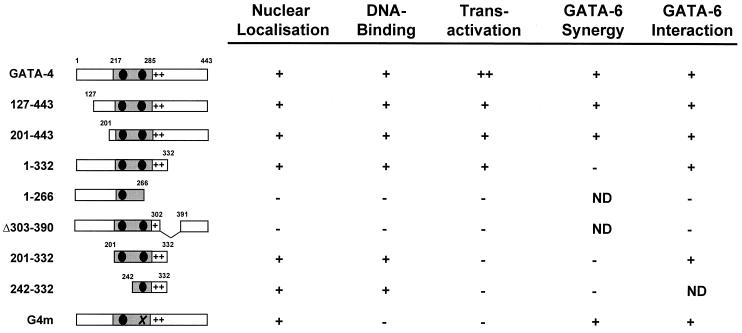
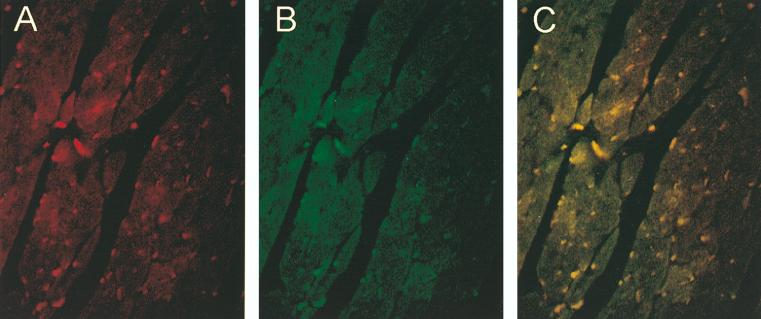
Two members of the GATA family of transcription factors, GATA-4 and GATA-6, are expressed in the developing and postnatal myocardium and are equally potent transactivators of several cardiac promoters. However, several in vitro and in vivo lines of evidence suggest distinct roles for the two factors in the heart. Since identification of the endogenous downstream targets of GATA factors would greatly help to elucidate their exact functions, we have developed an adenovirus-mediated antisense strategy to specifically inhibit GATA-4 and GATA-6 protein production in postnatal cardiomyocytes. Expression of several endogenous cardiac genes was significantly down-regulated in cells lacking GATA-4 or GATA-6, indicating that these factors are required for the maintenance of the cardiac genetic program. Interestingly, transcription of some genes like the alpha- and beta-myosin heavy-chain (alpha- and beta-MHC) genes was preferentially regulated by GATA-4 due, in part, to higher affinity of GATA-4 for their promoter GATA element. However, transcription of several other genes, including the atrial natriuretic factor and B-type natriuretic peptide (ANF and BNP) genes, was similarly down-regulated in cardiomyocytes lacking one or both GATA factors, suggesting that GATA-4 and GATA-6 could act through the same transcriptional pathway. Consistent with this, GATA-4 and GATA-6 were found to colocalize in postnatal cardiomyocytes and to interact functionally and physically to provide cooperative activation of the ANF and BNP promoters. The results identify for the first time bona fide in vivo targets for GATA-4 and GATA-6 in the myocardium. The data also show that GATA factors act in concert to regulate distinct subsets of genes, suggesting that combinatorial interactions among GATA factors may differentially control various cellular processes.
Figures
References
-
- Antakly T, Raquidan D, O’Donnel D, Katnick L. Regulation of glucocorticoid receptor expression. I. Use of a specific radioimmunoassay and antiserum to a synthetic peptide of the N-terminal domain. Endocrinology. 1990;126:1821–1828. - PubMed
-
- Argentin S, Sun Y-L, Lihrmann I, Schmidt T J, Drouin J, Nemer M. Distal cis-acting promoter sequences mediate glucocorticoid stimulation of cardiac atrial natriuretic factor gene transcription. J Biol Chem. 1991;266:23315–23322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials