

The role of beta7 integrins in CD8 T cell trafficking during an antiviral immune response
- PMID: 10330442
- PMCID: PMC2193647
- DOI: 10.1084/jem.189.10.1631
The role of beta7 integrins in CD8 T cell trafficking during an antiviral immune response
Erratum in
- J Exp Med 1999 Nov 1;190(9):following 1362
Abstract
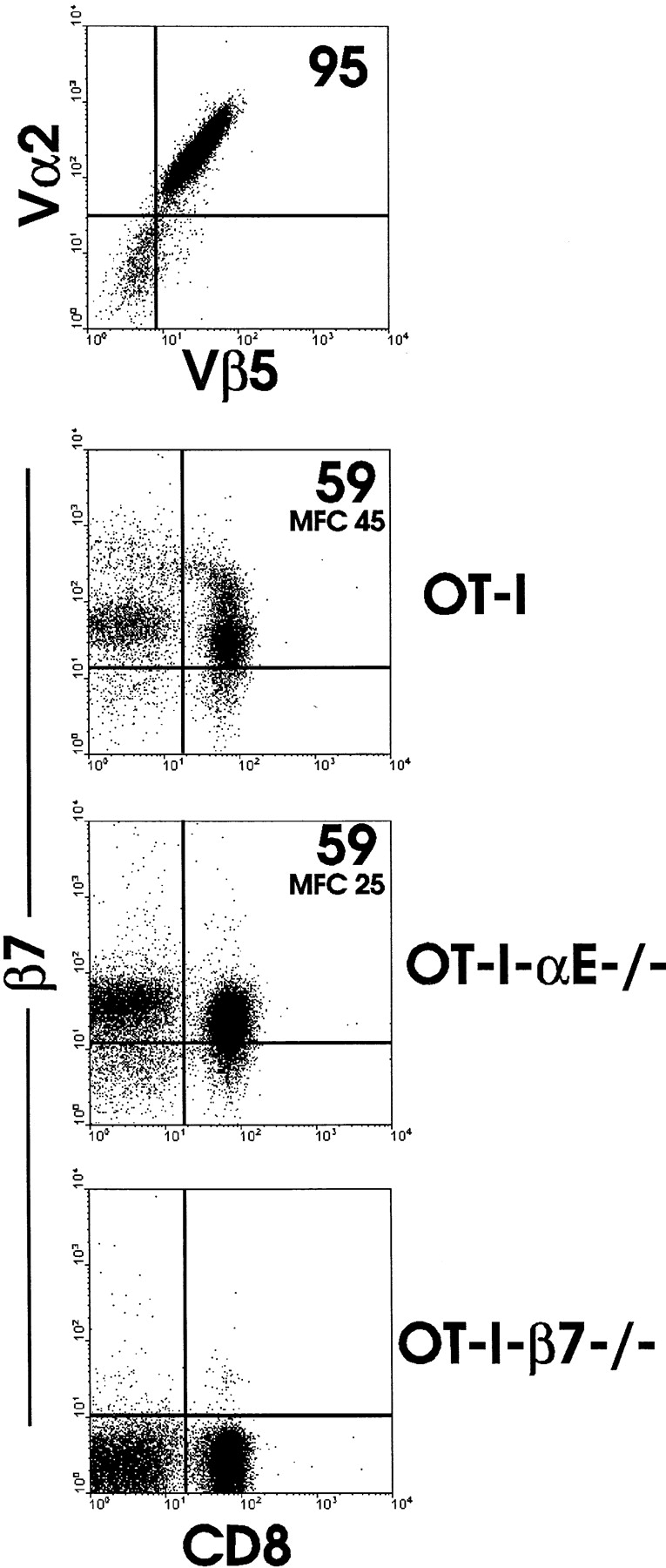
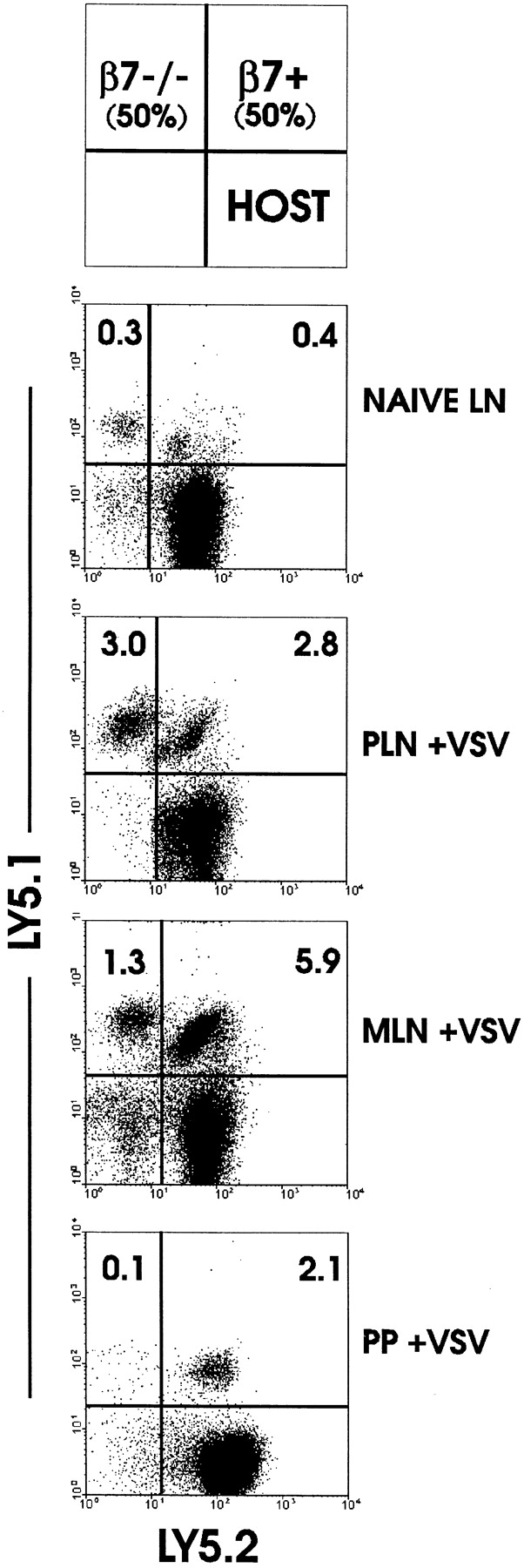
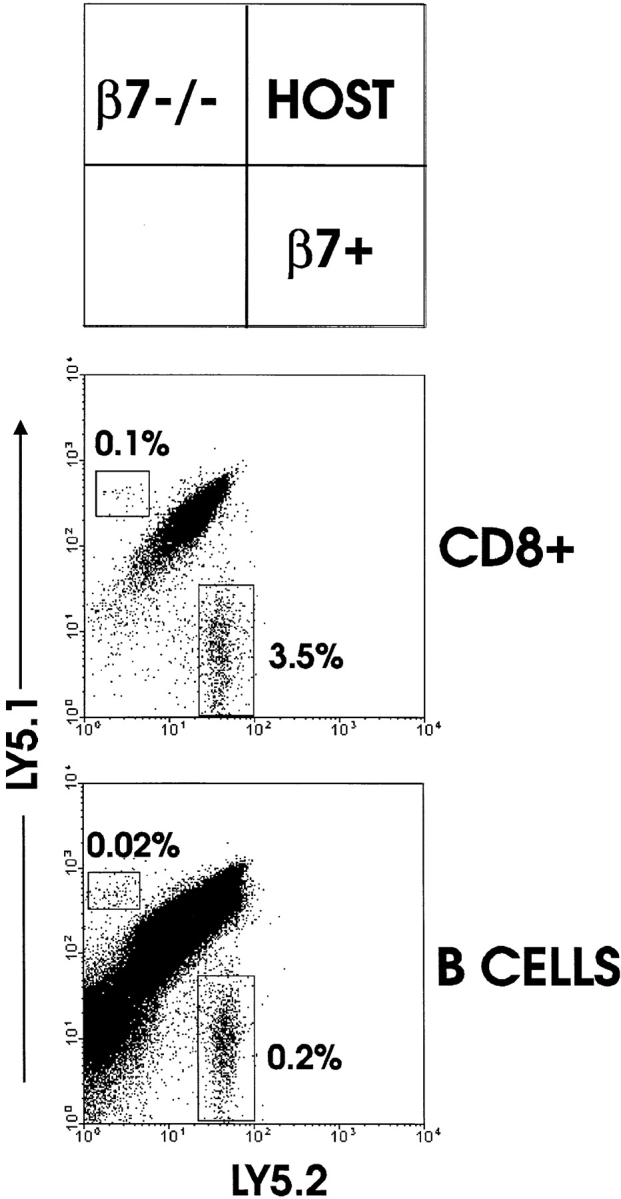
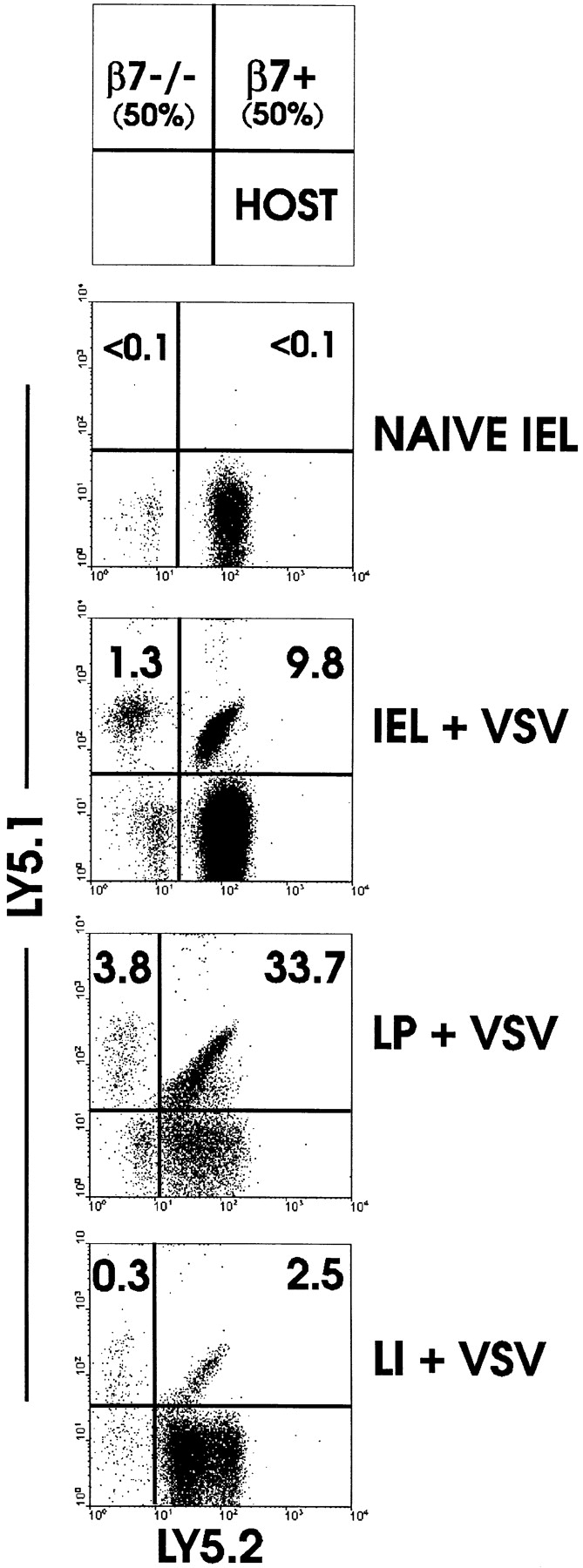
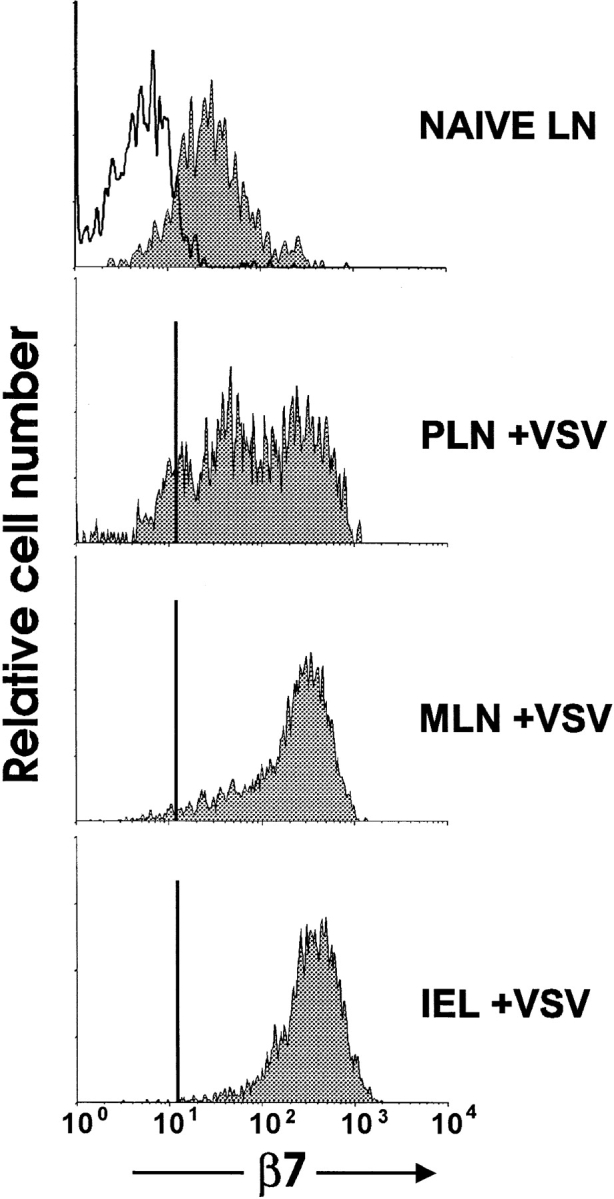
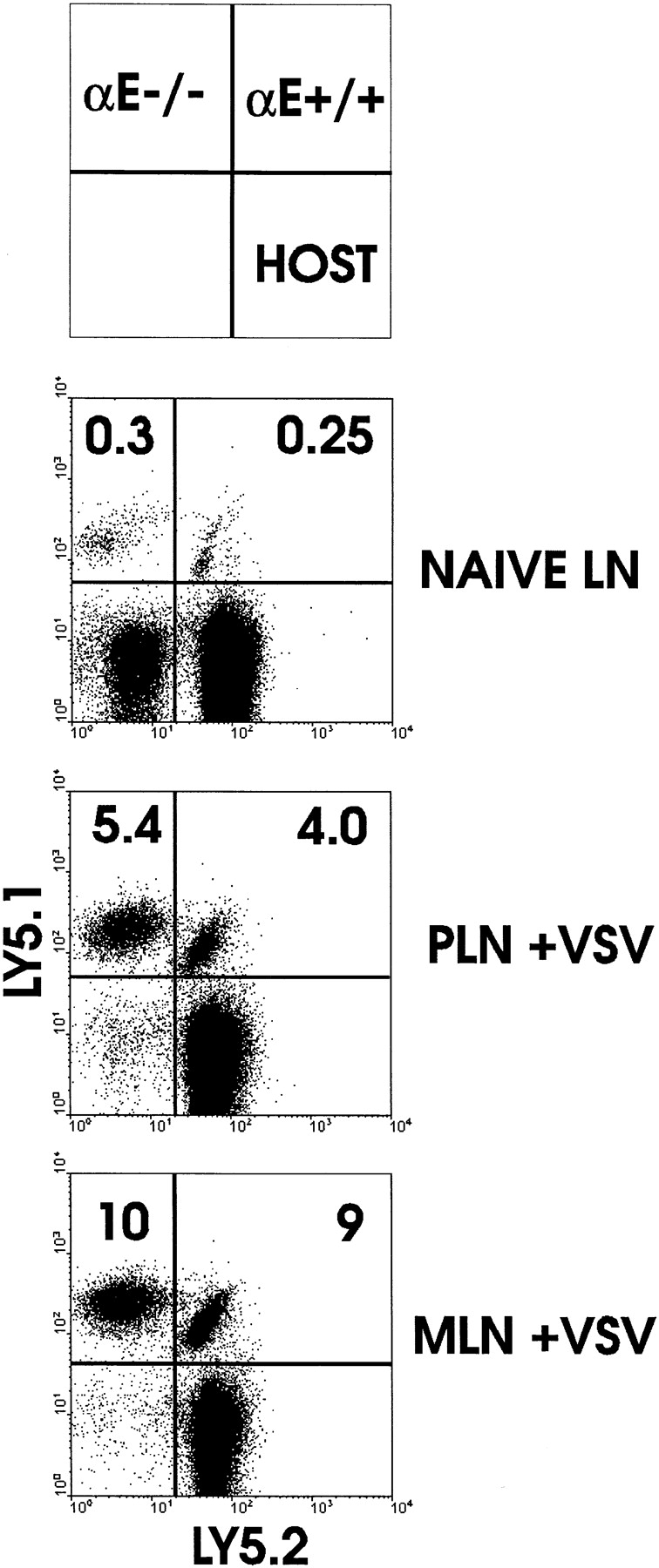
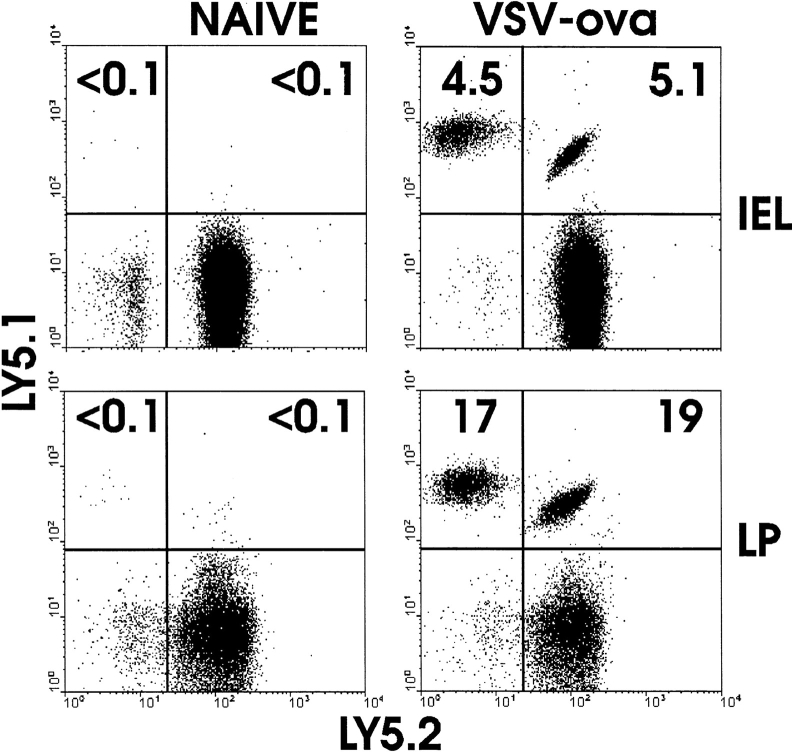
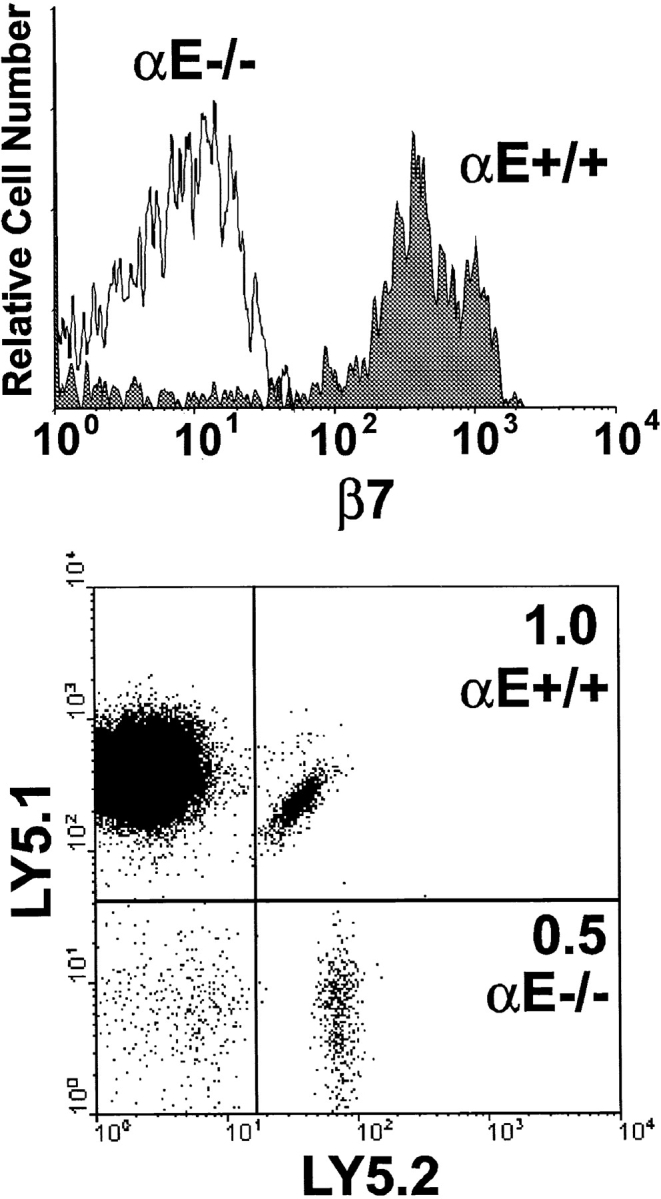
The requirement of beta7 integrins for lymphocyte migration was examined during an ongoing immune response in vivo. Transgenic mice (OT-I) expressing an ovalbumin-specific major histocompatibility complex class I-restricted T cell receptor for antigen were rendered deficient in expression of all beta7 integrins or only the alphaEbeta7 integrin. To quantitate the relative use of beta7 integrins in migration in vivo, equal numbers of OT-I and OT-I-beta7(-/-) or OT-I-alphaE-/- lymph node (LN) cells were adoptively transferred to normal mice. Although OT-I-beta7(-/-) LN cells migrated to mesenteric LN and peripheral LN as well as wild-type cells, beta7 integrins were required for naive CD8 T cell and B cell migration to Peyer's patch. After infection with a recombinant virus (vesicular stomatitis virus) encoding ovalbumin, beta7 integrins became critical for migration of activated CD8 T cells to the mesenteric LN and Peyer's patch. Naive CD8 T cells did not enter the lamina propria or the intestinal epithelium, and the majority of migration of activated CD8 T cells to the small and large intestinal mucosa, including the epithelium, was beta7 integrin-mediated. The alphaEbeta7 integrin appeared to play no role in migration during a primary CD8 T cell immune response in vivo. Furthermore, despite dramatic upregulation of alphaEbeta7 by CD8 T cells after entry into the epithelium, long-term retention of intestinal intraepithelial lymphocytes was also alphaEbeta7 independent.
Figures
References
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. - PubMed
-
- Garside P, Ingulli E, Merica RR, Johnson JG, Noelle RJ, Jenkins MK. Visualization of specific B and T lymphocyte interactions in the lymph node. Science. 1998;281:96–99. - PubMed
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76:301–314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
