

Suppression of platelet aggregation by Bordetella pertussis adenylate cyclase toxin
- PMID: 10338478
- PMCID: PMC96579
- DOI: 10.1128/IAI.67.6.2763-2768.1999
Suppression of platelet aggregation by Bordetella pertussis adenylate cyclase toxin
Abstract
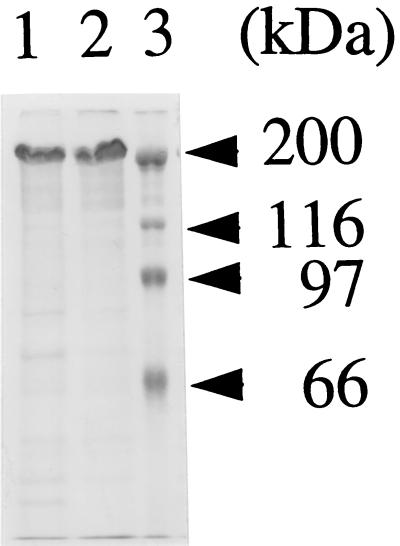
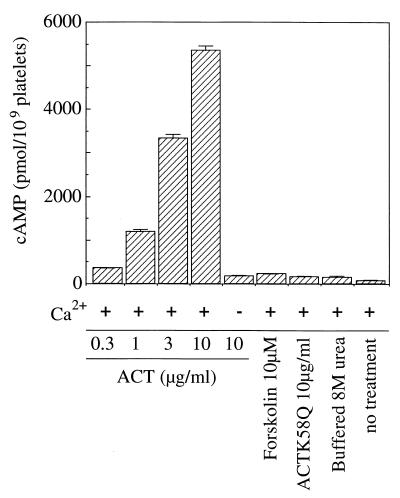
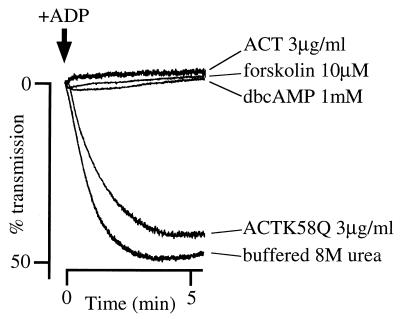
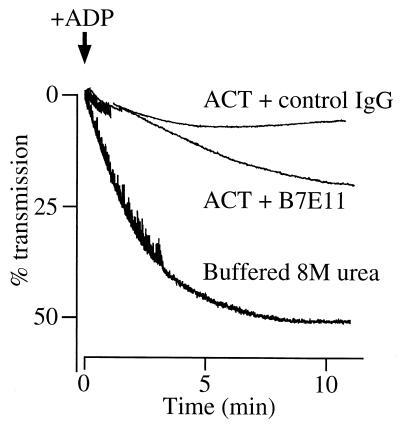
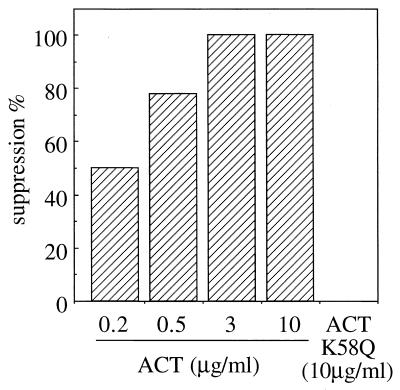
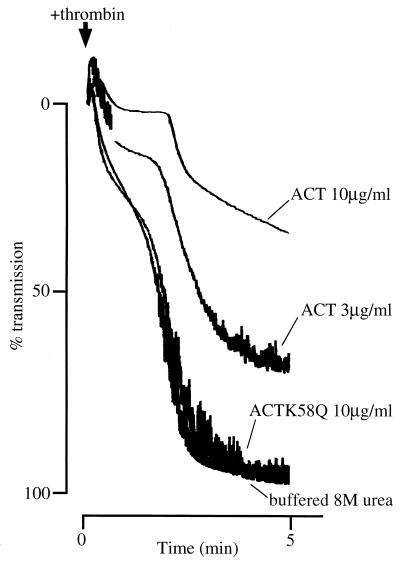
The effect of Bordetella pertussis adenylate cyclase toxin (ACT) on platelet aggregation was investigated. This cell-invasive adenylate cyclase completely suppressed ADP (10 microM)-induced aggregation of rabbit platelets at 3 micrograms/ml and strongly suppressed thrombin (0. 2 U/ml)-induced aggregation at 10 micrograms/ml. The suppression was accompanied by marked increase in platelet intracellular cyclic AMP (cAMP) content and was diminished by the anti-ACT monoclonal antibody B7E11. A catalytically inactive point mutant of ACT did not show the suppressive effect. Since an increase of cAMP content is a known cause of platelet dysfunction, these results indicate that the observed platelet inactivation was due to the catalytic activity of ACT through increase of intracellular cAMP.
Figures
References
-
- Benz R, Maier E, Ladant D, Ullmann A, Šebo P. Adenylate cyclase toxin of Bordetella pertussis: evidence for the formation of small ion-permeable channels and comparison with HlyA of Escherichia coli. J Biol Chem. 1994;269:27231–27239. - PubMed
-
- Born G V R. Aggregation of blood platelets by adenosine diphosphate and its reversal. Nature. 1962;194:927–929. - PubMed
-
- Brownlie R M, Coote J G, Parton R, Schultz J E, Rogel A, Hanski E. Cloning of the adenylate cyclase genetic determinant of Bordetella pertussis and its expression in Escherichia coli and B. pertussis. Microb Pathog. 1988;4:335–344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
