

Cell cycle-dependent regulation of RNA polymerase I transcription: the nucleolar transcription factor UBF is inactive in mitosis and early G1
- PMID: 10339547
- PMCID: PMC26841
- DOI: 10.1073/pnas.96.11.6096
Cell cycle-dependent regulation of RNA polymerase I transcription: the nucleolar transcription factor UBF is inactive in mitosis and early G1
Abstract
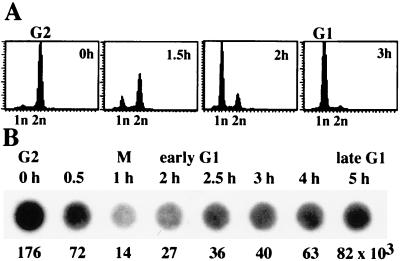
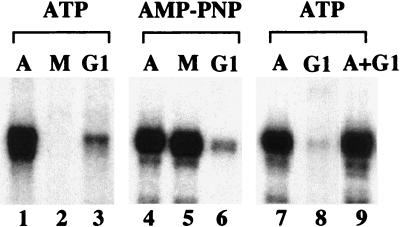
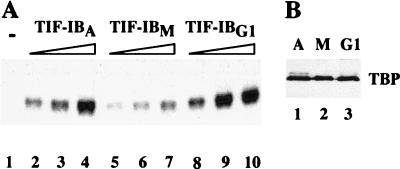
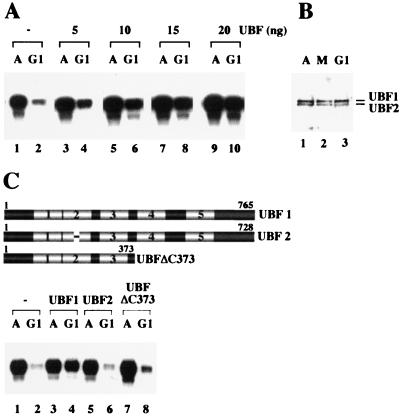
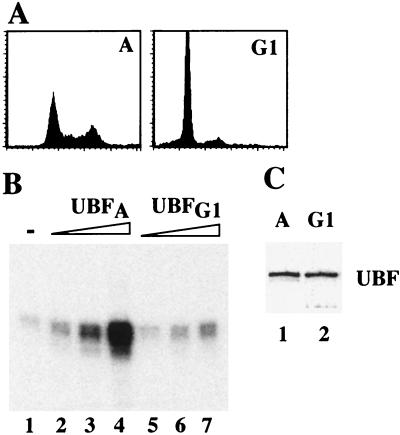
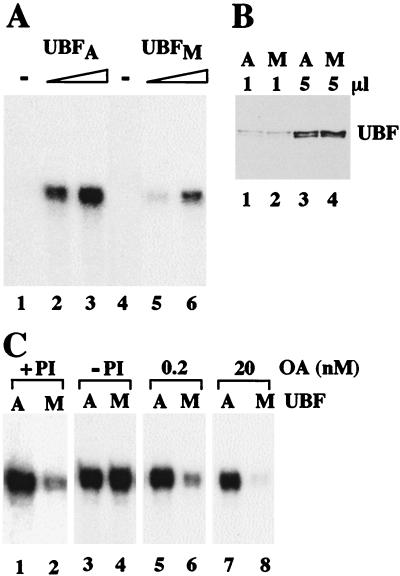
Transcription of ribosomal RNA genes by RNA polymerase (pol) I oscillates during the cell cycle, being maximal in S and G2 phase, repressed during mitosis, and gradually recovering during G1 progression. We have shown that transcription initiation factor (TIF)-IB/SL1 is inactivated during mitosis by cdc2/cyclin B-directed phosphorylation of TAFI110. In this study, we have monitored reactivation of transcription after exit from mitosis. We demonstrate that the pol I factor UBF is also inactivated by phosphorylation but recovers with different kinetics than TIF-IB/SL1. Whereas TIF-IB/SL1 activity is rapidly regained on entry into G1, UBF is reactivated later in G1, concomitant with the onset of pol I transcription. Repression of pol I transcription in mitosis and early G1 can be reproduced with either extracts from cells synchronized in M or G1 phase or with purified TIF-IB/SL1 and UBF isolated in the presence of phosphatase inhibitors. The results suggest that two basal transcription factors, e.g., TIF-IB/SL1 and UBF, are inactivated at mitosis and reactivated by dephosphorylation at the exit from mitosis and during G1 progression, respectively.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
