

doi: 10.1073/pnas.96.11.6107.
DNA methylation is a reversible biological signal
Affiliations
- PMID: 10339549
- PMCID: PMC26843
- DOI: 10.1073/pnas.96.11.6107
Item in Clipboard
DNA methylation is a reversible biological signal
Proc Natl Acad Sci U S A.
.
Abstract
The pattern of DNA methylation plays an important role in regulating different genome functions. To test the hypothesis that DNA methylation is a reversible biochemical process, we purified a DNA demethylase from human cells that catalyzes the cleavage of a methyl residue from 5-methyl cytosine and its release as methanol. We show that similar to DNA methyltransferase, DNA demethylase shows CpG dinucleotide specificity, can demethylate mdCpdG sites in different sequence contexts, and demethylates both fully methylated and hemimethylated DNA. Thus, contrary to the commonly accepted model, DNA methylation is a reversible signal, similar to other physiological biochemical modifications.
Figures
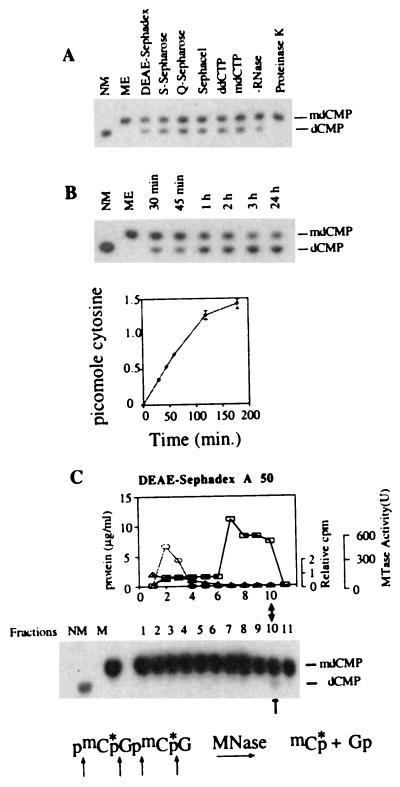
Purification of a dMTase from human cells. (A) [mdC32pdG]n double-stranded oligomer (1 ng) was incubated for 1 hour at 37°C with either buffer L (ME), 0.3 μg of active dMTase fractions from the different chromatography steps in the presence of 100 μg/ml RNase as indicated, with DEAE-Sephacel dMTase fraction in the presence of RNase, and either ddCTP (500 μM) or mdCTP (500 μM), with DEAE-Sephacel dMTase fraction in the absence of RNase, or with DEAE-Sephacel dMTase fraction after preincubation for 30 min. with proteinase K (200 μg/ml). The reaction products were digested to 3′ mononucleotides and analyzed on TLC (24). Methylated (ME) and nonmethylated (NM) [dC32pdG]n substrates were digested to 3′ mononucleotides and loaded on the TLC plate as controls. (B) Kinetics of dMTase activity. [mdC32pdG]n double-stranded oligomer (1 ng) was incubated with either DEAE-Sephacel dMTase fraction (0.3 μg) for different time points and analyzed for demethylation as above. A representative chromatogram is shown. Chromatograms were quantified by a PhosphorImager, and the rate of transformation of mdCMP to dCMP was calculated and plotted. The results are a summary of three independent determinations ± SD. (C) Nuclear extracts were fractionated on a DEAE-Sephadex column. Fifty-microliter samples from each fraction (total fraction volume was 500 μl) were incubated with 1 ng of [mdC32pdG]n double-stranded oligomer, digested to 3′ mononucleotides, and analyzed on TLC (24). Control methylated (ME) and nonmethylated (NM) [dC32pdG]n substrates were digested to 3′ mononucleotides and loaded on the TLC plate to indicate the expected position of dCMP and mdCMP. The active fraction is indicated by an arrow. DNA methyltransferase activity was determined by using a hemimethylated DNA substrate and [3H]AdoMet33 (○). One unit equals the amount of enzyme that catalyzes transfer of 1 pmol of tritiated methyl group on hemimethylated DNA per minute. The results are an average of three independent determinations. Protein concentration was determined by using the Bio-Rad Bradford kit (□). To test whether dCTP copurifies with dMTase, the elution profile of 20 μM of [α-32P]dCTP (1.1 × 106 dpm) incubated with the protein was determined by scintillation counting of the different DEAE fractions (▵) and presented as fraction of dCTP loaded on the column.
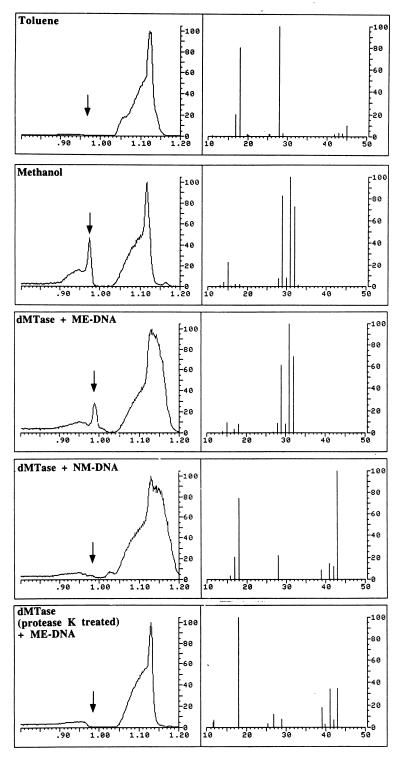
Purified dMTase releases the methyl group in the form of methanol. Shown are GC/MS analyses of reference materials and incubated samples for methanol. (Left) A portion of the gas chromatogram produced by the injected sample. The abscissa is the retention time in minutes and the ordinate is relative intensity in arbitrary units. (Right) The mass spectrum measured at the retention time of methanol (marked with an arrow, Left). The abscissa is the mass/charge (m/z) ratio for the ions detected, whereas the ordinate is the relative intensity of the ions detected. The large irregularly shaped peaks in the gas chromatograms are caused by hydrocarbon impurities in the toluene solvent that would normally be of no consequence and not detected except that the instrumental sensitivity required for methanol detection is such that they are also detected. “Toluene” illustrates the results of the GC/MS analysis of the toluene solvent used in this study. The mass spectrum recorded at the retention time for methanol does not present ions that are prominent in the mass spectrum for dilute authentic methanol in toluene solution (1:1,000 by volume, m/z 15, 29, 31, and 32, “Methanol”). “dMTase + ME DNA” illustrates the results obtained for the DNA demethylation reaction [with methylated CpG DNA (400 ng) and purified dMTase (4.8 μg) in a volume of 50 μl at 37°C overnight] and demonstrates that a peak with the retention time and mass spectrum consistent with its identification as methanol is detected. An additional ion at m/z 18 is most likely caused by residual water in the toluene extract. “dMTase + NM-DNA” and “dMTase (protease K treated) + ME-DNA” are the results of the analyses for demethylation reaction with CpG DNA with purified dMTase and protease K-treated dMTase with methylated CpG DNA [these reaction were performed with 4.8 μg of purified dMTase and 400 ng of DNA in a reaction volume of 50 μl with overnight incubation at 37°C, 10 units of protease K was used for treatment of 4.8 μg of dMTase in a reaction volume of 42 μl at 50°C for 3 hours], respectively, and display none of the ions associated with methanol. The ion at m/z 18 is because of residual water in the extract. Prominent ions in the range m/z 39–43 are because of trace hydrocarbons in the toluene.
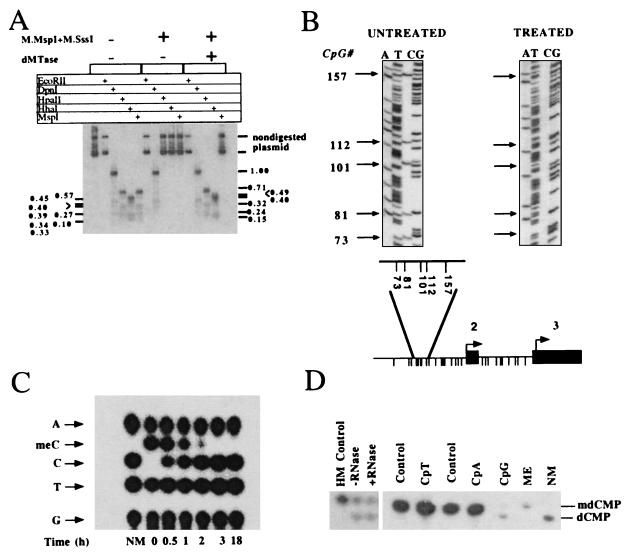
Substrate specificity of dMTase. (A) Demethylation of methylated SK plasmid. Plasmid pBluescript SK (5 ng) was methylated with SssI methylase and MspI methylase. dMTase-treated (1.2 μg of concentrated DEAE Sephacel fraction for 3 hours) (methylated SK + dMTase) and nontreated plasmid (methylated SK) as well as nonmethylated SK plasmid were digested with EcoRII, DpnI, HhaI, MspI, and HpaII. The digestion products were analyzed on a 2% agarose gel, blotted onto a Hybond N+ filter, and hybridized with a 32P-labeled SK probe. The expected HpaII/MspI fragments are indicated on the right side of the figure, the expected HhaI fragments are indicated on the left side of the figure. Note that control plasmids are not digested with MspI because the plasmid is methylated with MspI methyltransferase). (B) Bisulfite methylation analysis of dMTase-treated pMetCAT plasmid. pMetCAT plasmid (24), methylated to completion with SssI, was subjected to a dMTase reaction as above. The dMTase-treated plasmid and nontreated control were subjected to bisulfite treatment as described (31), and the genomic region bearing five CpG sites upstream of the second exon of DNA methyltransferase was amplified by using PCR (31) and sequenced. Twenty clones were sequenced per DNA sample. Unmethylated cytosines are converted to thymidines by this protocol, whereas methylated cytosines are protected and are visualized as cytosines. One representative sequencing gel of bisulfite treated DNA is presented per condition. Arrows indicate the specific CpG sites by their position. The numbering is according to GenBank accession no. M84387. (Lower) The physical map of the genomic region residing upstream of exon 2 of DNA methyltransferase (indicated by filled boxes). Intronic sequence is indicated by a line. A blowup of the region amplified after bisulfite treatment is shown above the physical map. The different CpG sites in the fragment are presented as descending lines. Under these conditions, all the clones of the demethylase-treated plasmids were demethylated. (C) Plasmid pBluescript SK (5 ng) was methylated with SssI methylase and MspI methylase. The methylated pBluescript SK was used as template to DNA synthesis by using DNA polymerase and [α32-P]dGTP, mdCTP, dTTP, dATP, and hexanucleotide primers. After two rounds of purification on NAP-5 column, the substrate was subjected to demethylation by using 0.12 μg of purified dMTase for different time intervals as indicated. (D) Demethylation of hemimethylated [mdC32pdG]n·[dCpdG]n (HM), [mdC32pdA]n (CpA), and [mdC32pdT]n (CpT) DNA substrates (15). One nanogram of the indicated substrates, as well as a double methylated substrate [mdC32pdG]n (CpG), were incubated either in the presence (0.3 μg) or absence of dMTase (control for CpT and CpA and ME for CpG) and were then digested to 3′ mononucleotides and analyzed by TLC. Hemimethylated DNA was incubated with dMTase in the presence or absence of RNase (100 μg/ml). Nonmethylated [dC32pdG]n served as a control for the position of dCMP.
Comment in
-
DNA demethylation.Proc Natl Acad Sci U S A. 1999 May 25;96(11):5894-6. doi: 10.1073/pnas.96.11.5894. Proc Natl Acad Sci U S A. 1999. PMID: 10339513 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
