

Regulation of RpoS proteolysis in Escherichia coli: the response regulator RssB is a recognition factor that interacts with the turnover element in RpoS
- PMID: 10339606
- PMCID: PMC26900
- DOI: 10.1073/pnas.96.11.6439
Regulation of RpoS proteolysis in Escherichia coli: the response regulator RssB is a recognition factor that interacts with the turnover element in RpoS
Abstract
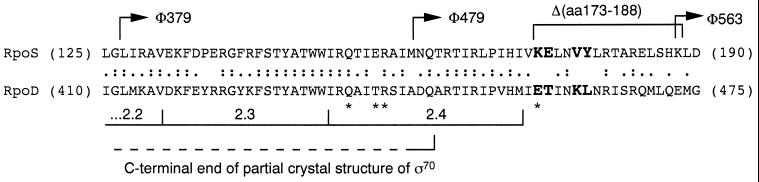
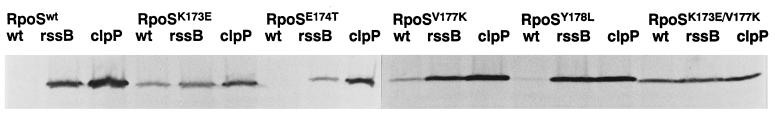
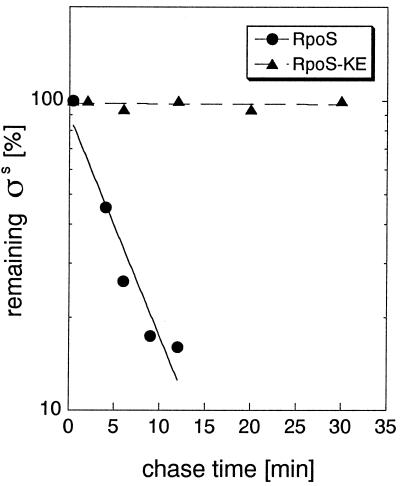
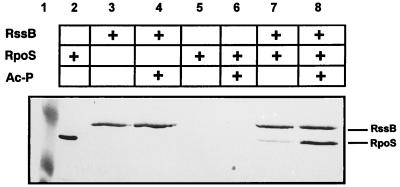
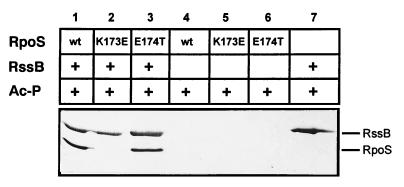
The degradation of the RpoS (sigmaS) subunit of RNA polymerase in Escherichia coli is a prime example of regulated proteolysis in prokaryotes. RpoS turnover depends on ClpXP protease, the response regulator RssB, and a hitherto uncharacterized "turnover element" within RpoS itself. Here we localize the turnover element to a small element (around the crucial amino acid lysine-173) directly downstream of the promoter-recognizing region 2.4 in RpoS. Its sequence as well as its location identify the turnover element as a unique proteolysis-promoting motif. This element is shown to be a site of interaction with RssB. Thus, RssB is functionally unique among response regulators as a direct recognition factor in ClpXP-dependent RpoS proteolysis. Binding of RssB to RpoS is stimulated by phosphorylation of the RssB receiver domain, suggesting that environmental stress affects RpoS proteolysis by modulating RssB affinity for RpoS. Initial evidence indicates that lysine-173 in RpoS, besides being essential of RpoS proteolysis, may play a role in promoter recognition. Thus the same region in RpoS is crucial for proteolysis as well as for activity as a transcription factor.
Figures
References
-
- Hengge-Aronis R. In: Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology. Neidhardt F C, editor. Washington, DC: Am. Soc. Microbiol.; 1996. pp. 1497–1512.
-
- Loewen P C, Hengge-Aronis R. Annu Rev Microbiol. 1994;48:53–80. - PubMed
-
- Lange R, Hengge-Aronis R. Genes Dev. 1994;8:1600–1612. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
