

Electron microscopic evidence against apoptosis as the mechanism of neuronal death in global ischemia
- PMID: 10341224
- PMCID: PMC6782610
- DOI: 10.1523/JNEUROSCI.19-11-04200.1999
Electron microscopic evidence against apoptosis as the mechanism of neuronal death in global ischemia
Abstract
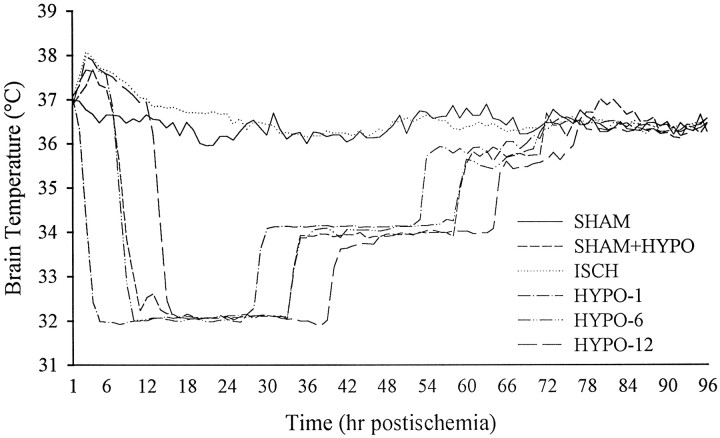
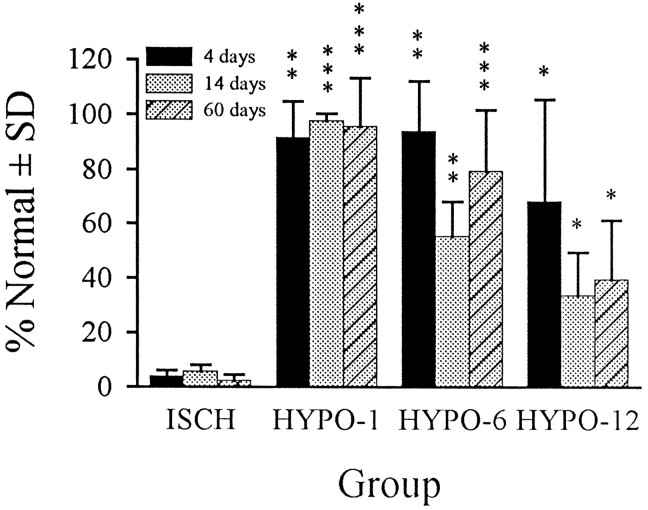
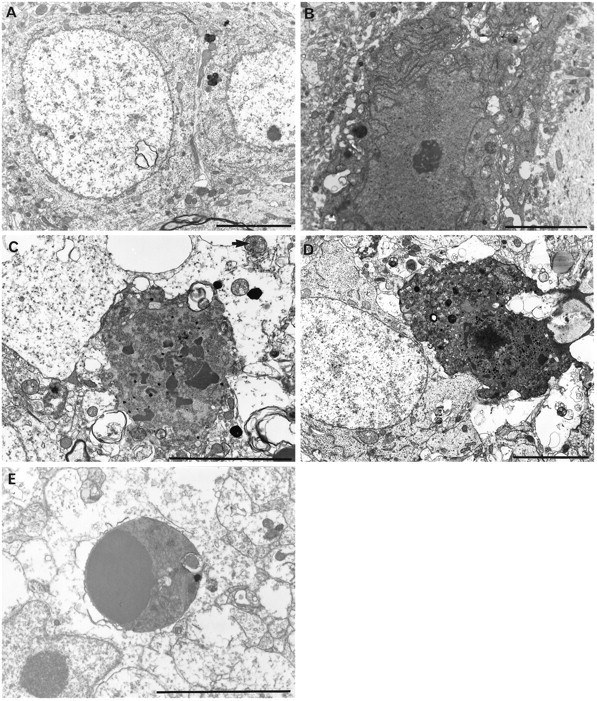
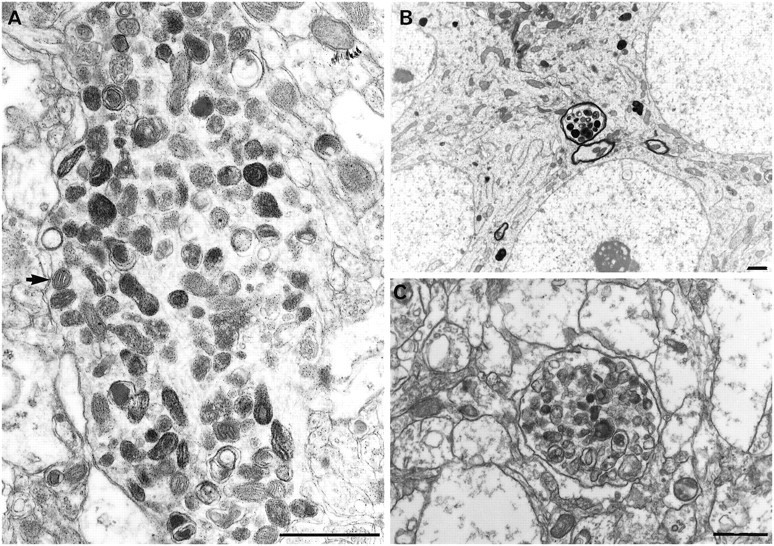
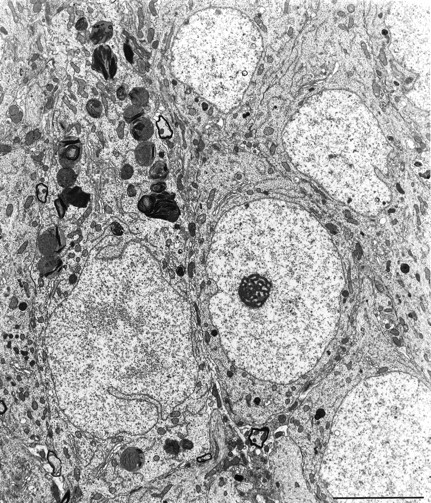
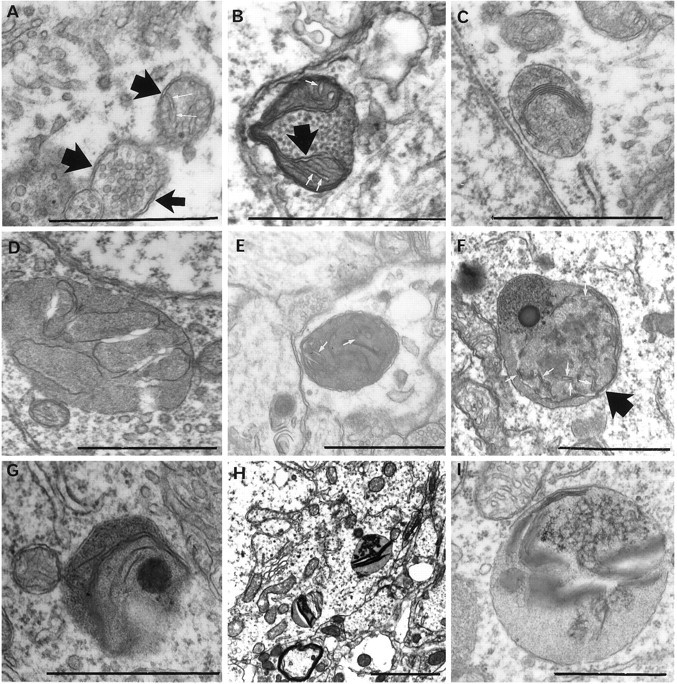
It has been repeatedly claimed that neuronal death in the hippocampal CA1 sector after untreated global ischemia occurs via apoptosis. This is based largely on DNA laddering, nick end labeling, and light microscopy. Delineation of apoptosis requires fine structural examination to detect morphological events of cell death. We studied the light and ultrastructural characteristics of CA1 injury after 5 min of untreated global ischemia in gerbils. To increase the likelihood of apoptosis, some ischemic gerbils were subjected to delayed postischemic hypothermia, a treatment that mitigates injury and delays the death of some neurons. In these gerbils, 2 d of mild hypothermia was initiated 1, 6, or 12 hr after ischemia, and gerbils were killed 4, 14, or 60 d later. Ischemia without subsequent cooling killed 96% of CA1 neurons by day 4, whereas all hypothermia-treated groups had significantly reduced injury at all survival times (2-67% loss). Electron microscopy of ischemic neurons with or without postischemic hypothermia revealed features of necrotic, not apoptotic, neuronal death even in cells that died 2 months after ischemia. Dilated organelles and intranuclear vacuoles preceded necrosis. Unique to the hypothermia-treated ischemic groups, some salvaged neurons were persistently abnormal and showed accumulation of unusual, morphologically complex secondary lysosomes. These indicate selective mitochondrial injury, because they were closely associated with normal and degenerate mitochondria, and transitional forms between mitochondria and lysosomes occurred. The results show that untreated global ischemic injury has necrotic, not apoptotic, morphology but do not rule out programmed biochemical events of the apoptotic pathway occurring before neuronal necrosis.
Figures



References
-
- Abe K, Aoki M, Kawagoe J, Yoshida T, Hattori A, Kogure K, Itoyama Y. Ischemic delayed neuronal death. A mitochondrial hypothesis. Stroke. 1995;26:1478–1489. - PubMed
-
- Auer RN. Structural neurotoxicologic investigation of the glycine antagonist 5-nitro-6,7-dichloroquinoxalinedione (ACEA1021). Neurotoxicology. 1997;18:53–62. - PubMed
-
- Bicknell GR, Cohen GM. Cleavage of DNA to large kilobase pair fragments occurs in some forms of necrosis as well as apoptosis. Biochem Biophys Res Commun. 1995;207:40–47. - PubMed
-
- Bonnekoh P, Barbier A, Oschlies U, Hossmann KA. Selective vulnerability in the gerbil hippocampus: morphological changes after 5-min ischemia and long survival times. Acta Neuropathol (Berl) 1990;80:18–25. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous