

Neuroprotection at Drosophila synapses conferred by prior heat shock
- PMID: 10341239
- PMCID: PMC6782588
- DOI: 10.1523/JNEUROSCI.19-11-04360.1999
Neuroprotection at Drosophila synapses conferred by prior heat shock
Abstract
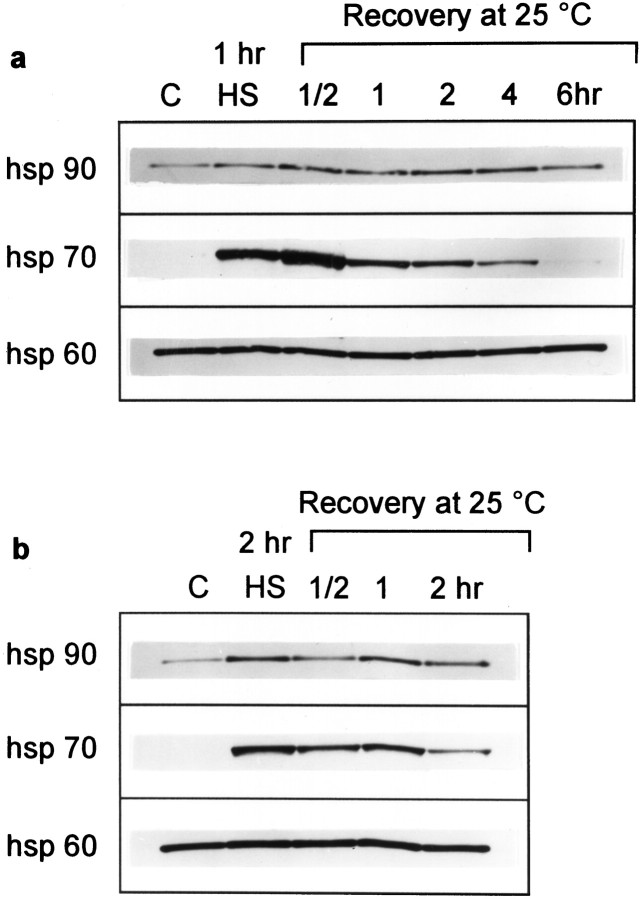
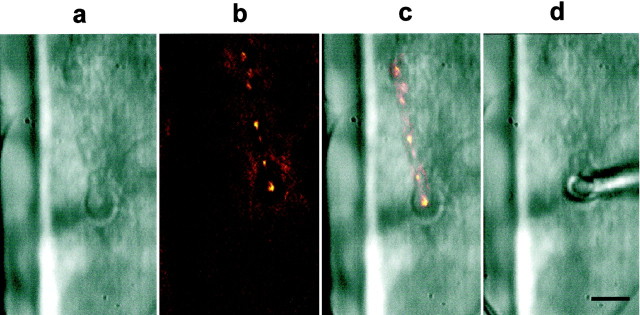
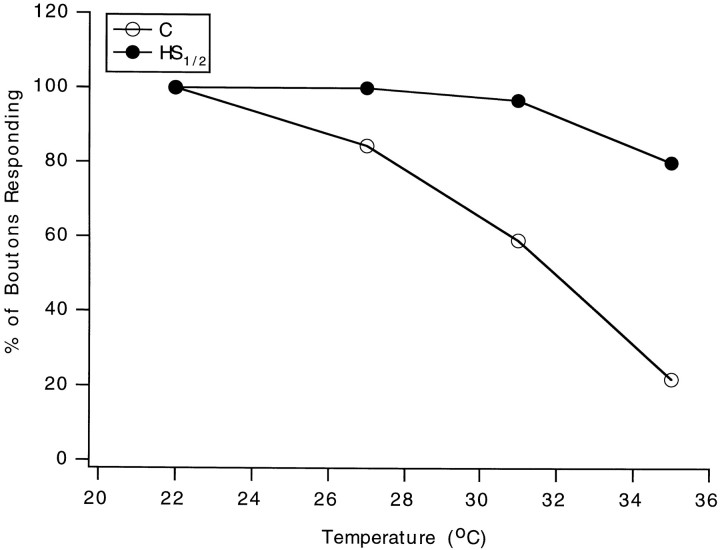
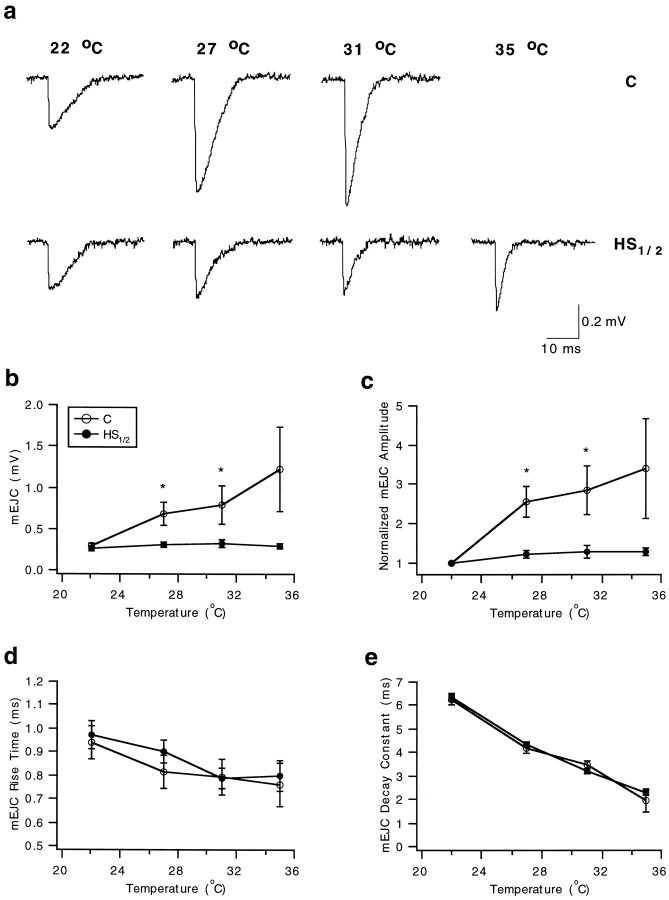
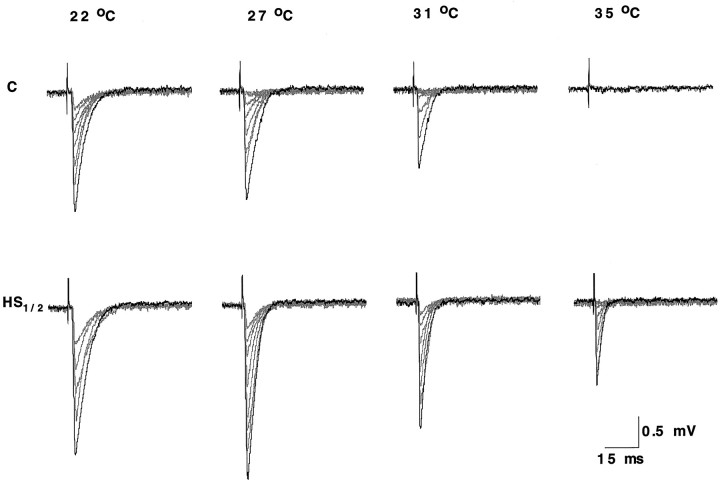
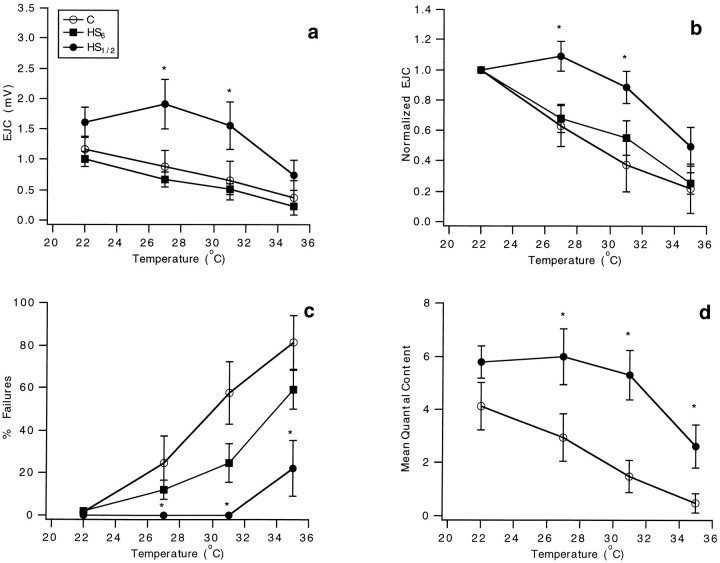
Synapses are critical sites of information transfer in the nervous system, and it is important that their functionality be maintained under stressful conditions to prevent communication breakdown. Here we show that synaptic transmission at the Drosophila larval neuromuscular junction is protected by prior exposure to heat shock that strongly induces expression of heat shock proteins, in particular hsp70. Using a macropatch electrode to record synaptic activity at individual, visualized boutons, we found that prior heat shock sustains synaptic performance at high test temperatures through pre- and postsynaptic alterations. After heat shock, nerve impulses release more quantal units at high temperatures and exhibit fewer failures of release (presynaptic modification), whereas the amplitude of quantal currents remains more constant than does that in nonheat-shocked controls (postsynaptic modification). The time course of these physiological changes is similar to that of elevated hsp70. Thus, stress-induced neuroprotective mechanisms maintain function at synapses by modifying their properties.
Figures
Similar articles
-
Enhancement of presynaptic performance in transgenic Drosophila overexpressing heat shock protein Hsp70.Synapse. 2002 Apr;44(1):8-14. doi: 10.1002/syn.10048. Synapse. 2002. PMID: 11842441
-
Role for calcium in heat shock-mediated synaptic thermoprotection in Drosophila larvae.J Neurobiol. 2003 Sep 15;56(4):360-71. doi: 10.1002/neu.10247. J Neurobiol. 2003. PMID: 12918020
-
Synaptic thermoprotection in a desert-dwelling Drosophila species.J Neurobiol. 2005 Aug;64(2):170-80. doi: 10.1002/neu.20132. J Neurobiol. 2005. PMID: 15818554
-
Glutamate receptors in synaptic assembly and plasticity: case studies on fly NMJs.Adv Exp Med Biol. 2012;970:3-28. doi: 10.1007/978-3-7091-0932-8_1. Adv Exp Med Biol. 2012. PMID: 22351049 Review.
-
[Regulation of heat shock gene expression in response to stress].Mol Biol (Mosk). 2017 May-Jun;51(3):400-417. doi: 10.7868/S0026898417020100. Mol Biol (Mosk). 2017. PMID: 28707656 Review. Russian.
Cited by
-
Induction of heat shock proteins in differentiated human neuronal cells following co-application of celastrol and arimoclomol.Cell Stress Chaperones. 2016 Sep;21(5):837-48. doi: 10.1007/s12192-016-0708-2. Epub 2016 Jun 8. Cell Stress Chaperones. 2016. PMID: 27273088 Free PMC article.
-
Expression of the Hsp23 chaperone during Drosophila embryogenesis: association to distinct neural and glial lineages.BMC Dev Biol. 2003 Nov 14;3:9. doi: 10.1186/1471-213X-3-9. BMC Dev Biol. 2003. PMID: 14617383 Free PMC article.
-
Temperature dependent plasticity of habituation in the crayfish.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011 Nov;197(11):1073-81. doi: 10.1007/s00359-011-0668-z. Epub 2011 Jul 23. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011. PMID: 21789652
-
Behavioral and electrophysiological analysis of general anesthesia in 3 background strains of Drosophila melanogaster.Fly (Austin). 2015;9(1):7-15. doi: 10.1080/19336934.2015.1072663. Epub 2015 Aug 12. Fly (Austin). 2015. PMID: 26267354 Free PMC article.
-
Quantal size and variation determined by vesicle size in normal and mutant Drosophila glutamatergic synapses.J Neurosci. 2002 Dec 1;22(23):10267-76. doi: 10.1523/JNEUROSCI.22-23-10267.2002. J Neurosci. 2002. PMID: 12451127 Free PMC article.
References
-
- Akner G, Mossberg K, Sundqvist KG, Gustafsson JA, Wikstrom AC. Evidence for reversible non-microtubule and non-microfilament dependent nuclear translocation of hsp90 after heat shock in human fibroblasts. Eur J Cell Biol. 1992;58:356–364. - PubMed
-
- Atwood HL, Govind CK, Wu C-F. Differential ultrastructure of synaptic terminals on ventral longitudinal abdominal muscles in Drosophila larvae. J Neurobiol. 1993;24:1008–1024. - PubMed
-
- Barbe MF, Tytell M, Gower DJ, Welch WJ. Hyperthermia protects against light damage in the rat retina. Science. 1988;241:1817–1820. - PubMed
-
- Brand AH, Manoukian AS, Perrimon N. Ectopic gene expression in Drosophila. Methods Cell Biol. 1994;44:635–654. - PubMed
-
- Chamberlain LH, Burgoyne RD. The molecular chaperone function of the secretory vesicle cysteine string proteins. J Biol Chem. 1997a;272:31420–31426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases