

Sensory feedback can coordinate the swimming activity of the leech
- PMID: 10341261
- PMCID: PMC6782618
- DOI: 10.1523/JNEUROSCI.19-11-04634.1999
Sensory feedback can coordinate the swimming activity of the leech
Abstract
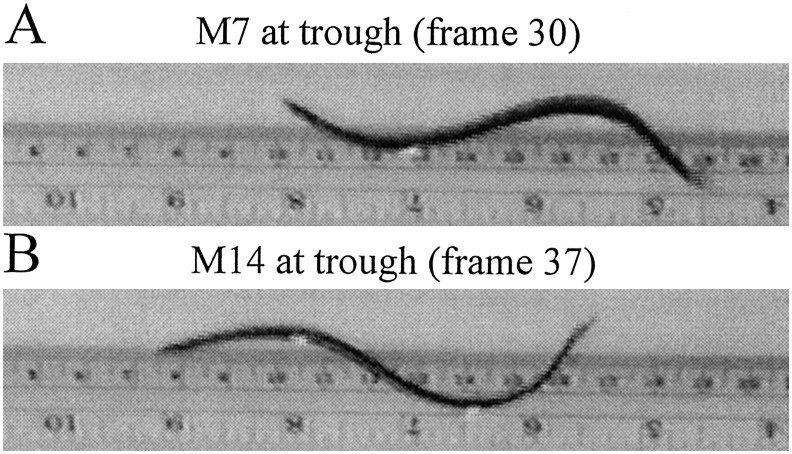
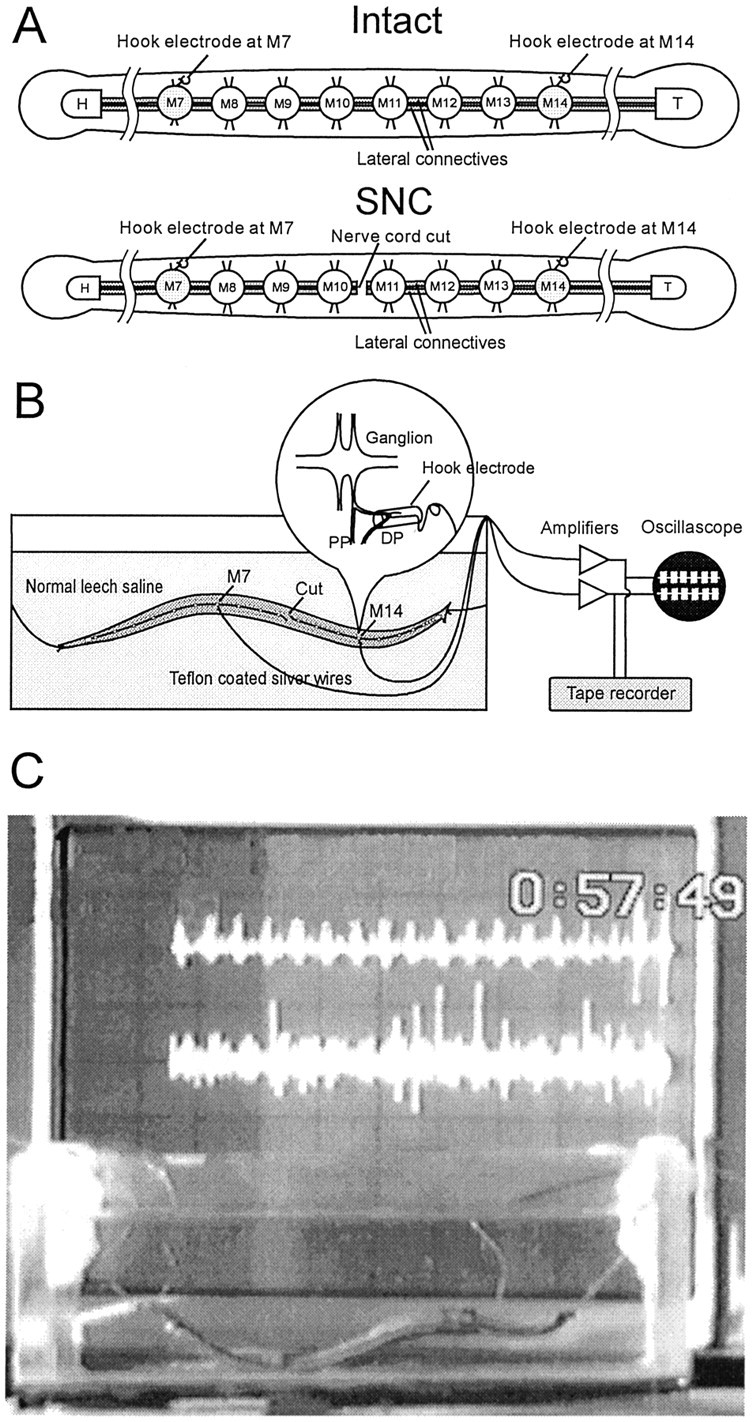
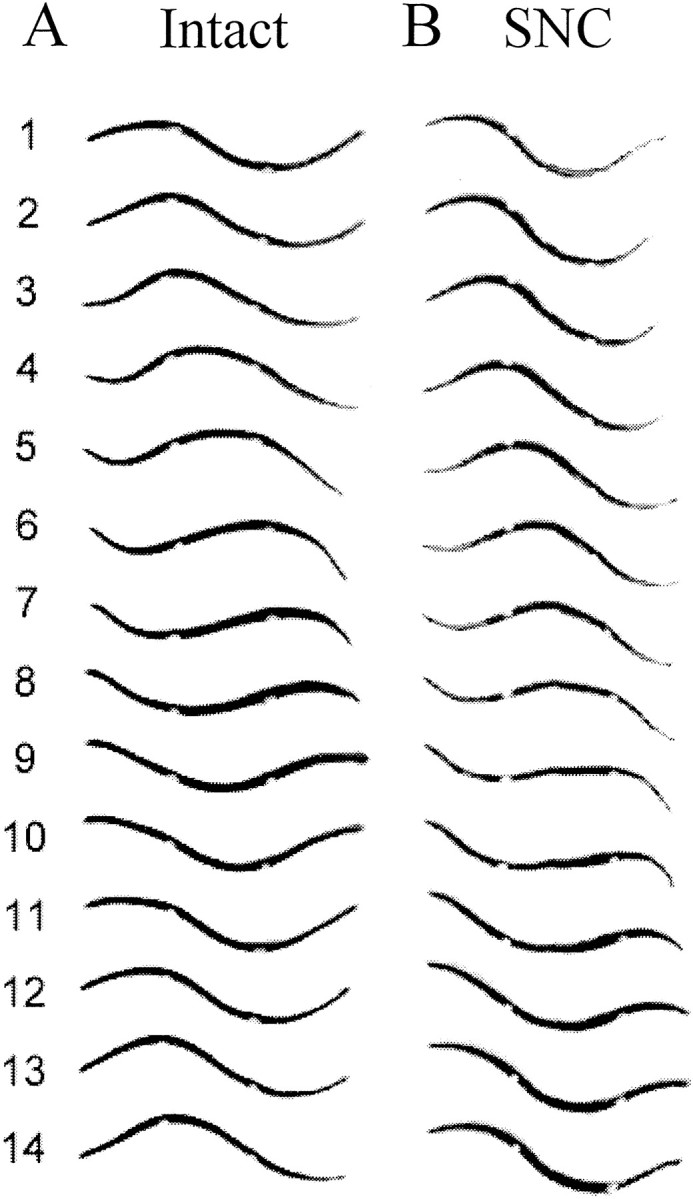
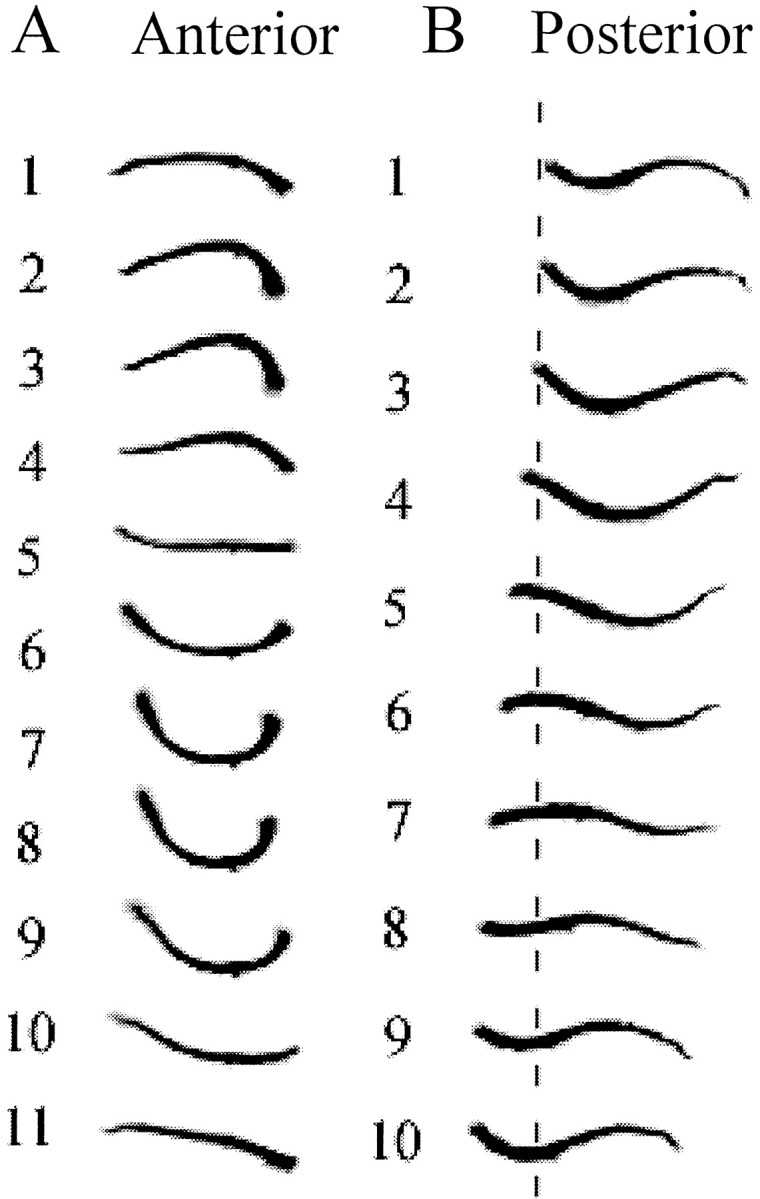
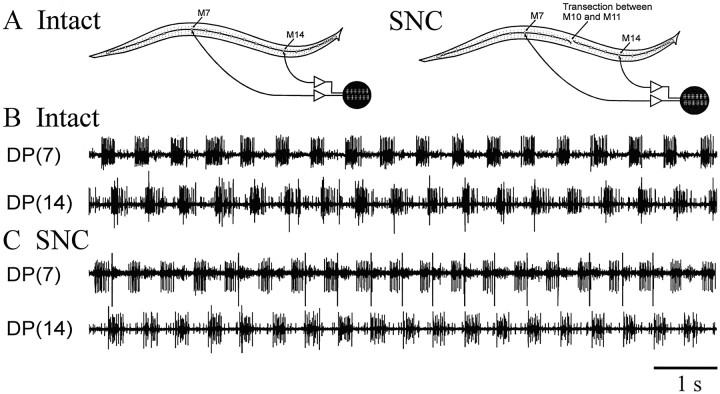
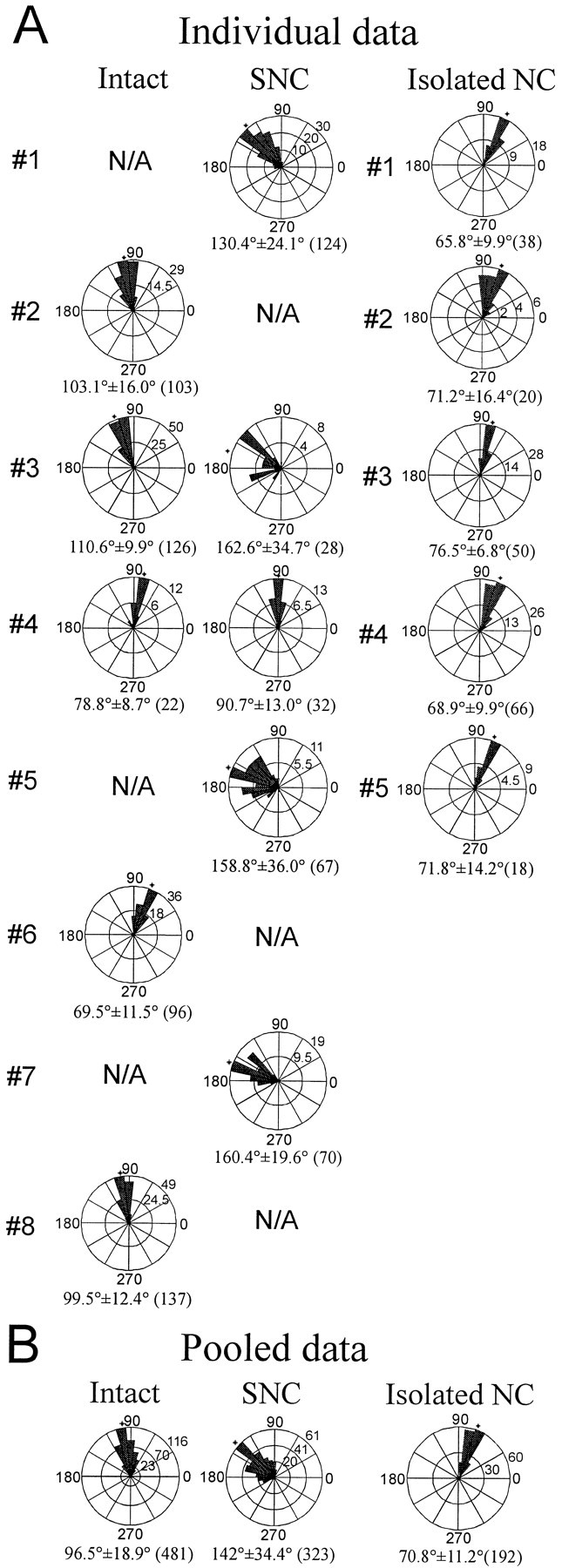
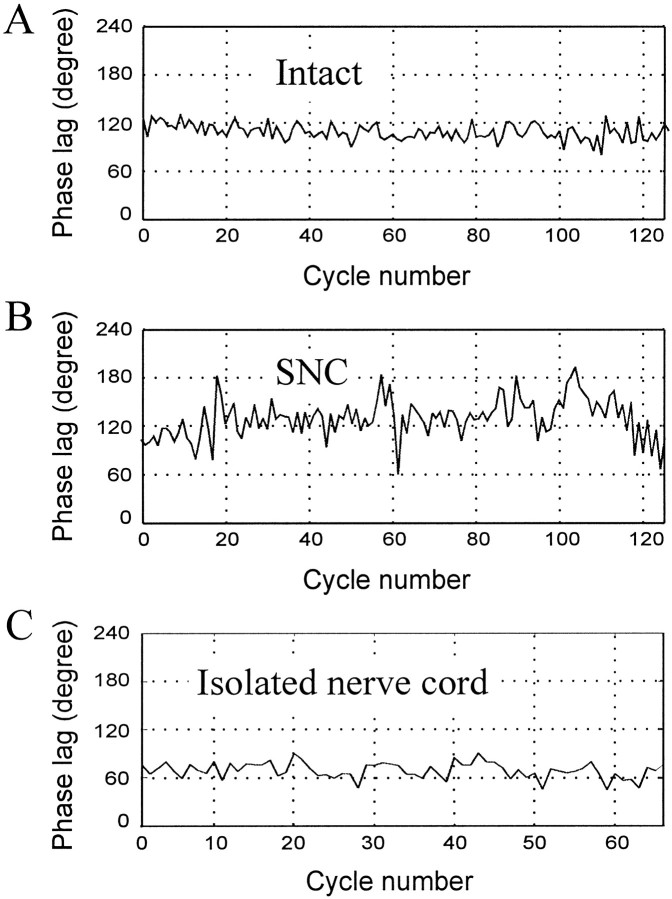
Previous studies showed that sensory feedback from the body wall is important and sometimes critical for generating normal, robust swimming activity in leeches. In this paper, we evaluate the role of sensory feedback in intersegmental coordination using both behavioral and physiological measurements. We severed the ventral nerve cord of leeches in midbody and then made video and in situ extracellular recordings from swimming animals. Our electrophysiological recordings unequivocally demonstrate that active intersegmental coordination occurs in leeches with severed nerve cords, refuting earlier conclusions that sensory feedback cannot coordinate swimming activity. Intersegmental coordination can in fact be achieved by sensory feedback alone, without the intersegmental interactions conveyed by the nerve cord.
Figures

Similar articles
-
Neuronal control of swimming behavior: comparison of vertebrate and invertebrate model systems.Prog Neurobiol. 2011 Feb;93(2):244-69. doi: 10.1016/j.pneurobio.2010.11.001. Epub 2010 Nov 18. Prog Neurobiol. 2011. PMID: 21093529 Free PMC article. Review.
-
Model for intersegmental coordination of leech swimming: central and sensory mechanisms.J Neurophysiol. 2002 Jun;87(6):2760-9. doi: 10.1152/jn.2002.87.6.2760. J Neurophysiol. 2002. PMID: 12037178
-
Intersegmental coordination of leech swimming: comparison of in situ and isolated nerve cord activity with body wall movement.Brain Res. 1984 May 14;299(2):363-6. doi: 10.1016/0006-8993(84)90720-0. Brain Res. 1984. PMID: 6733455
-
Entrainment of leech swimming activity by the ventral stretch receptor.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004 Nov;190(11):939-49. doi: 10.1007/s00359-004-0549-9. Epub 2004 Aug 25. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004. PMID: 15338181
-
Neuronal control of leech swimming.J Neurobiol. 1995 Jul;27(3):403-18. doi: 10.1002/neu.480270312. J Neurobiol. 1995. PMID: 7673898 Review.
Cited by
-
Positive feedback loops sustain repeating bursts in neuronal circuits.J Biol Phys. 2011 Jun;37(3):317-45. doi: 10.1007/s10867-010-9210-8. Epub 2010 Dec 16. J Biol Phys. 2011. PMID: 22654180 Free PMC article.
-
Compensatory plasticity restores locomotion after chronic removal of descending projections.J Neurophysiol. 2015 Jun 1;113(10):3610-22. doi: 10.1152/jn.00135.2015. Epub 2015 Mar 18. J Neurophysiol. 2015. PMID: 25787951 Free PMC article.
-
Mechanisms underlying rhythmic locomotion: dynamics of muscle activation.J Exp Biol. 2011 Jun 1;214(Pt 11):1955-64. doi: 10.1242/jeb.052787. J Exp Biol. 2011. PMID: 21562183 Free PMC article.
-
Neuronal control of swimming behavior: comparison of vertebrate and invertebrate model systems.Prog Neurobiol. 2011 Feb;93(2):244-69. doi: 10.1016/j.pneurobio.2010.11.001. Epub 2010 Nov 18. Prog Neurobiol. 2011. PMID: 21093529 Free PMC article. Review.
-
Formal analysis of resonance entrainment by central pattern generator.J Math Biol. 2008 Aug;57(2):183-207. doi: 10.1007/s00285-007-0151-1. Epub 2008 Jan 4. J Math Biol. 2008. PMID: 18175118
References
-
- Bassler U. The walking- (and searching-) pattern generator of stick insects, a modular system composed of reflex chains and endogenous oscillators. Biol Cybern. 1993;69:305–317.
-
- Brodfuehrer PD, Friesen WO. Control of leech swimming activity by cephalic ganglia. J Neurobiol. 1986;17:697–705. - PubMed
-
- Cohen AH, Wallen P. The neuronal correlate of locomotion in fish: “fictive swimming” induced in an in vitro preparation of the lamprey spinal cord. Exp Brain Res. 1980;41:11–18. - PubMed
-
- Delcomyn F. Neural basis of rhythmic behavior in animals. Science. 1980;210:492–498. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources