

Path integration absent in scent-tracking fimbria-fornix rats: evidence for hippocampal involvement in "sense of direction" and "sense of distance" using self-movement cues
- PMID: 10341264
- PMCID: PMC6782623
- DOI: 10.1523/JNEUROSCI.19-11-04662.1999
Path integration absent in scent-tracking fimbria-fornix rats: evidence for hippocampal involvement in "sense of direction" and "sense of distance" using self-movement cues
Abstract
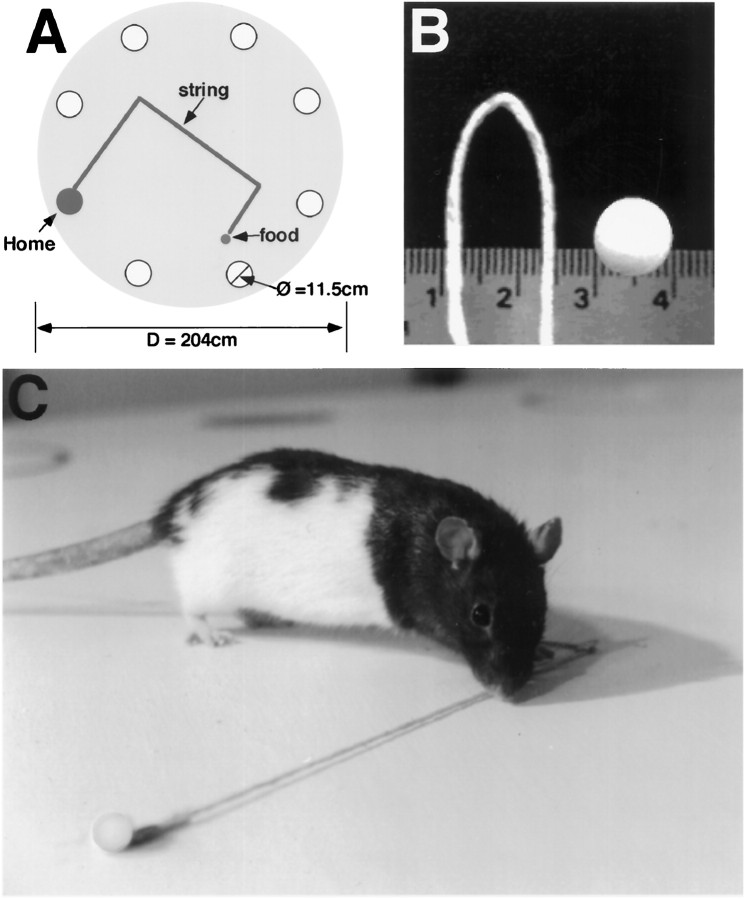
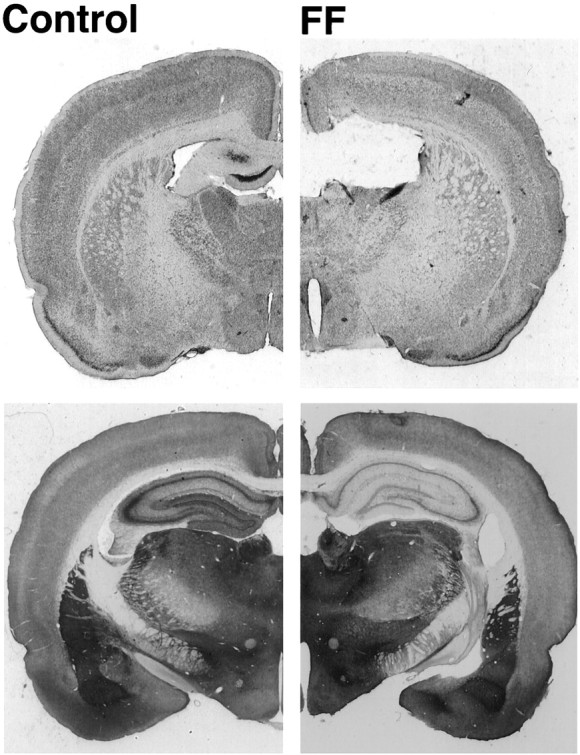
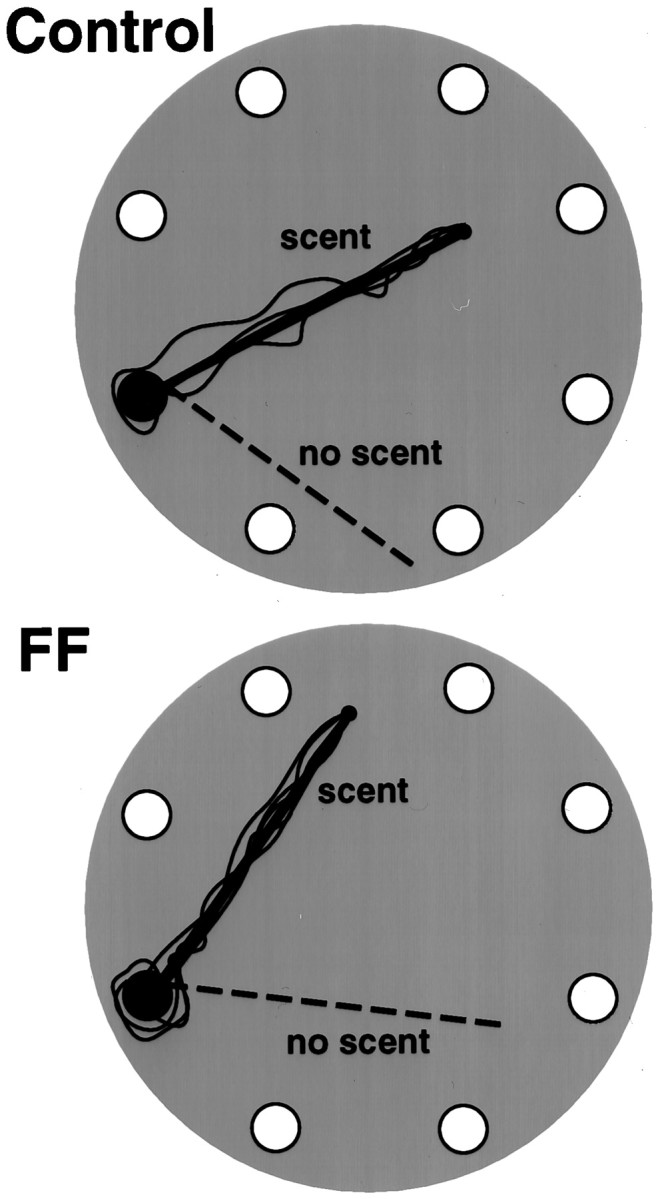
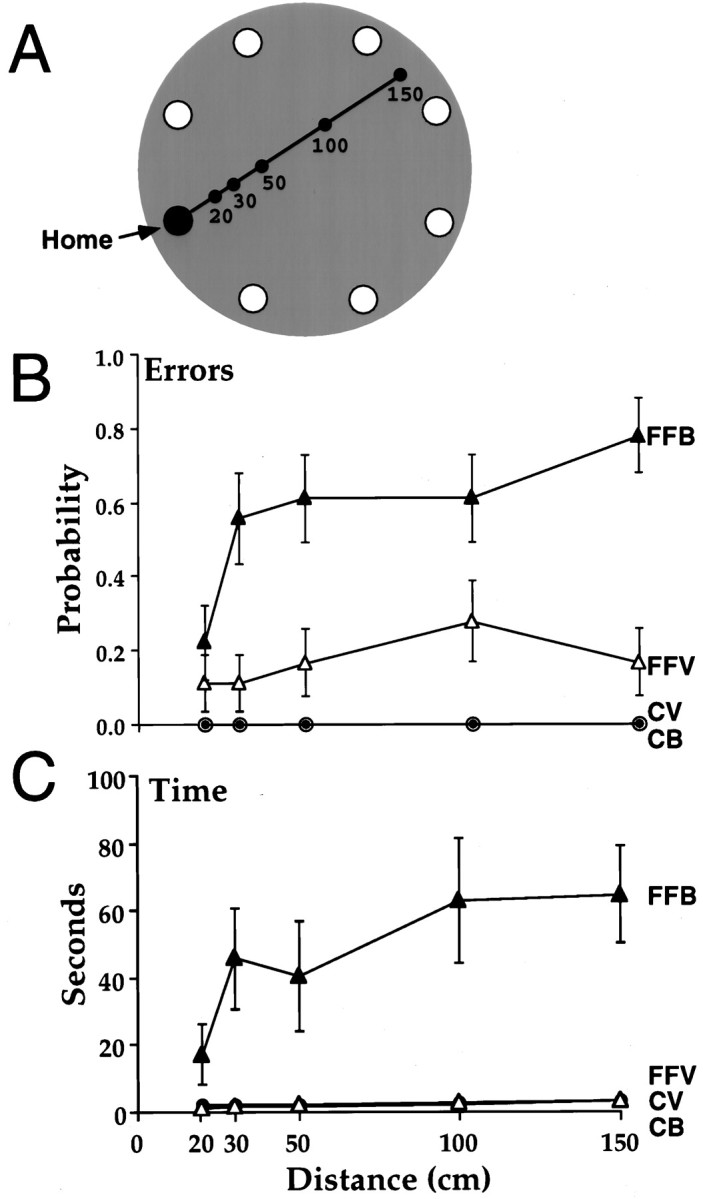
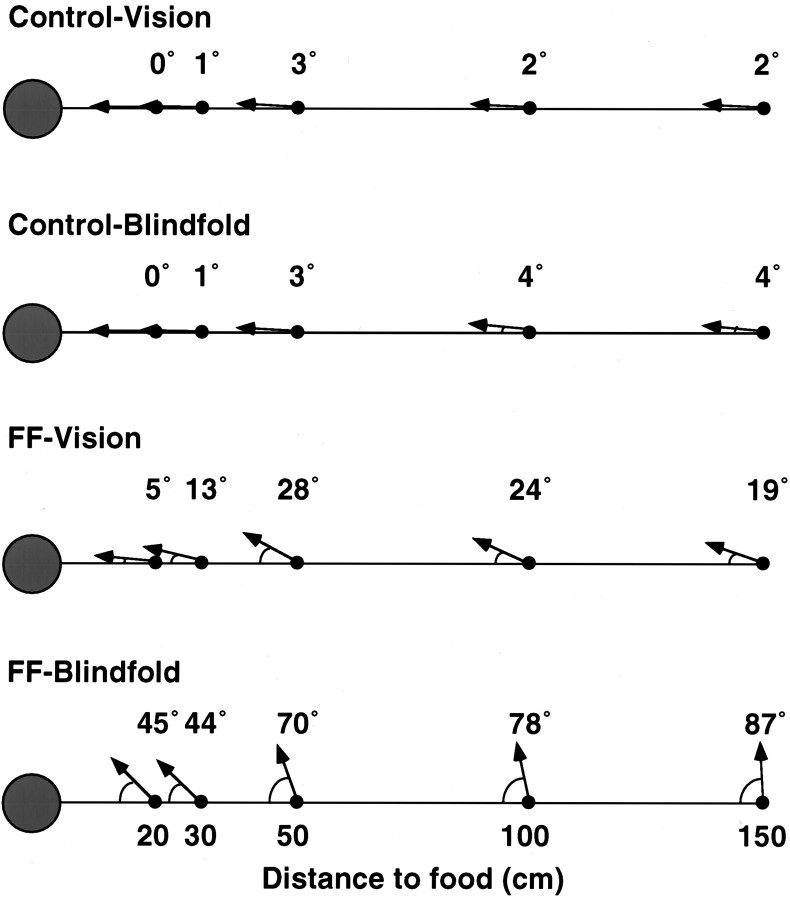
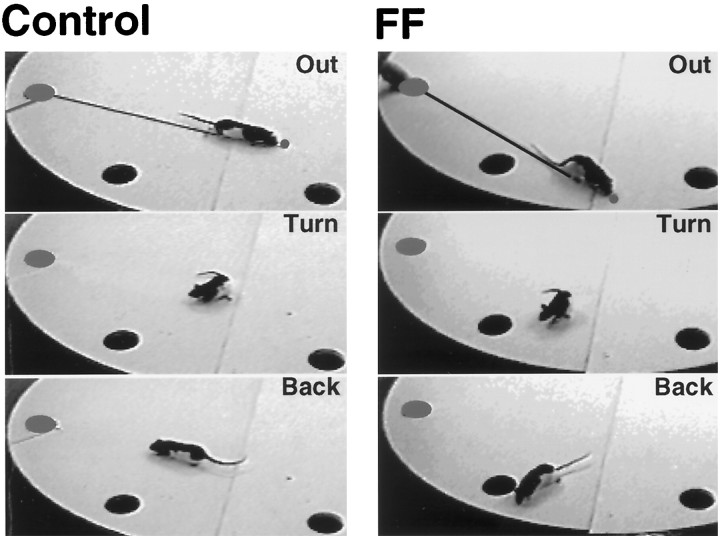
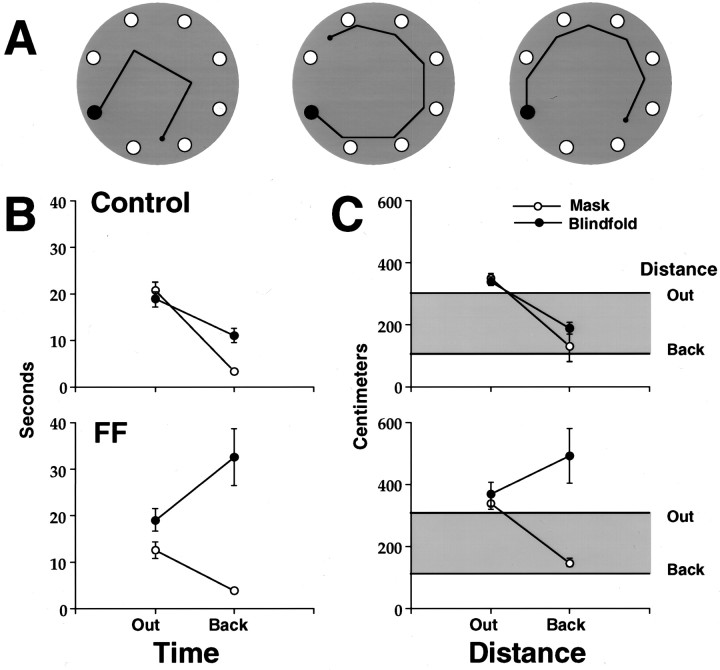
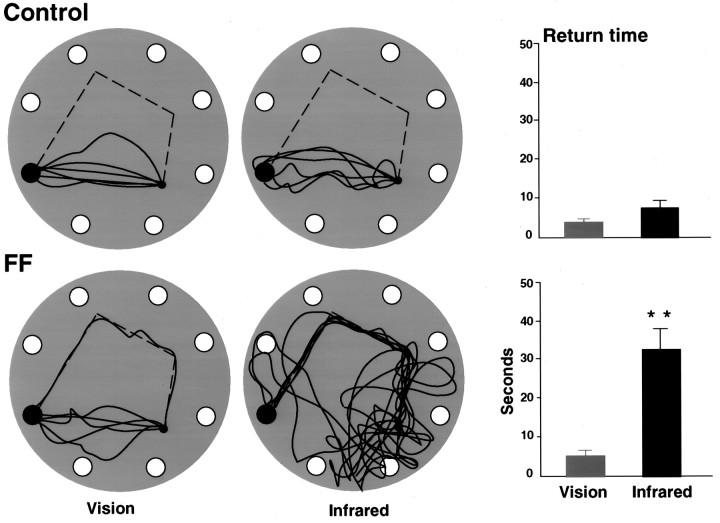
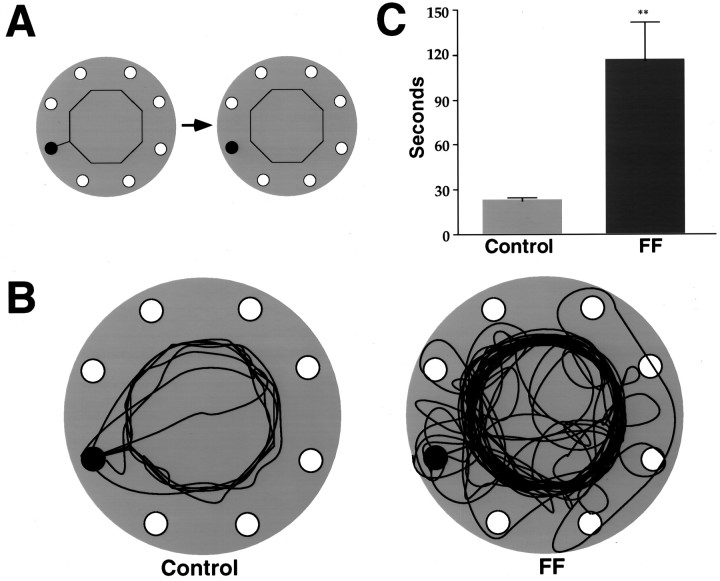
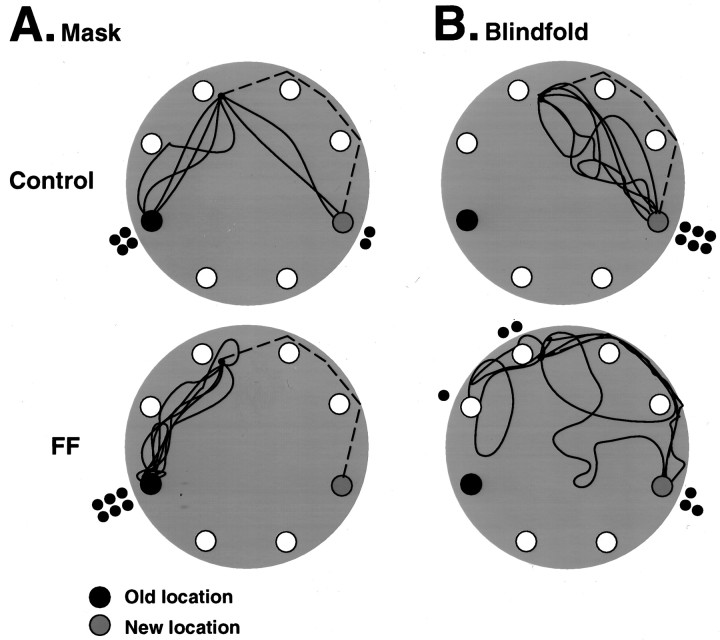
Allothetic and idiothetic navigation strategies use very different cue constellations and computational processes. Allothetic navigation requires the use of the relationships between relatively stable external (visual, olfactory, auditory) cues, whereas idiothetic navigation requires the integration of cues generated by self-movement and/or efferent copy of movement commands. The flexibility with which animals can switch between these strategies and the neural structures that support these strategies are not well understood. By capitalizing on the proclivity of foraging rats to carry large food pellets back to a refuge for eating, the present study examined the contribution of the hippocampus to the use of allothetic versus idiothetic navigation strategies. Control rats and fimbria-fornix-ablated rats were trained to follow linear, polygonal, and octagonal scent trails that led to a piece of food. The ability of the rats to return to the refuge with the food via the shortest route using allothetic cues (visual cues and/or the odor trail available) or using ideothetic cues (the odor trail removed and the rats blindfolded or tested in infrared light) was examined. Control rats "closed the polygon" by returning directly home in all cue conditions. Fimbria-fornix rats successfully used allothetic cues (closed the polygon using visual cues or tracked back on the string) but were insensitive to the direction and distance of the refuge and were lost when restricted to idiothetic cues. The results support the hypothesis that the hippocampal formation is necessary for navigation requiring the integration of idiothetic cues.
Figures
Similar articles
-
Calibrating space: exploration is important for allothetic and idiothetic navigation.Hippocampus. 1999;9(6):659-67. doi: 10.1002/(SICI)1098-1063(1999)9:6<659::AID-HIPO7>3.0.CO;2-E. Hippocampus. 1999. PMID: 10641759
-
Dead reckoning (path integration) requires the hippocampal formation: evidence from spontaneous exploration and spatial learning tasks in light (allothetic) and dark (idiothetic) tests.Behav Brain Res. 2001 Dec 14;127(1-2):49-69. doi: 10.1016/s0166-4328(01)00359-x. Behav Brain Res. 2001. PMID: 11718884
-
A video demonstration of preserved piloting by scent tracking but impaired dead reckoning after fimbria-fornix lesions in the rat.J Vis Exp. 2009 Apr 24;(26):1193. doi: 10.3791/1193. J Vis Exp. 2009. PMID: 19398947 Free PMC article.
-
The Neurobiology of Mammalian Navigation.Curr Biol. 2018 Sep 10;28(17):R1023-R1042. doi: 10.1016/j.cub.2018.05.050. Curr Biol. 2018. PMID: 30205053 Review.
-
Hippocampal lesions and path integration.Curr Opin Neurobiol. 1997 Apr;7(2):228-34. doi: 10.1016/s0959-4388(97)80011-6. Curr Opin Neurobiol. 1997. PMID: 9142750 Review.
Cited by
-
Speed and Oscillations: Medial Septum Integration of Attention and Navigation.Front Syst Neurosci. 2017 Sep 20;11:67. doi: 10.3389/fnsys.2017.00067. eCollection 2017. Front Syst Neurosci. 2017. PMID: 28979196 Free PMC article. Review.
-
Vestibular information is required for dead reckoning in the rat.J Neurosci. 2002 Nov 15;22(22):10009-17. doi: 10.1523/JNEUROSCI.22-22-10009.2002. J Neurosci. 2002. PMID: 12427858 Free PMC article.
-
Temporary inactivation of the retrosplenial cortex causes a transient reorganization of spatial coding in the hippocampus.J Neurosci. 2001 Jun 1;21(11):3986-4001. doi: 10.1523/JNEUROSCI.21-11-03986.2001. J Neurosci. 2001. PMID: 11356886 Free PMC article.
-
Otolithic information is required for homing in the mouse.Hippocampus. 2015 Aug;25(8):890-9. doi: 10.1002/hipo.22410. Epub 2015 Jan 20. Hippocampus. 2015. PMID: 25565056 Free PMC article.
-
Hippocampal spatial representations require vestibular input.Hippocampus. 2002;12(3):291-303. doi: 10.1002/hipo.1112. Hippocampus. 2002. PMID: 12099481 Free PMC article.
References
-
- Amaral DG, Witter MP. Hippocampal formation. In: Paxinos G, editor. The rat nervous system. Academic; San Diego: 1995. pp. 443–493.
-
- Angeli SJ, Murray EA, Mishkin M. Hippocampectomized monkeys can remember one place but not two. Neuropsychologia. 1993;31:1021–1030. - PubMed
-
- Barlow JS. Inertial navigation as a basis for animal navigation. J Theor Biol. 1964;6:76–117. - PubMed
-
- Barnes CA. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol. 1979;93:74–104. - PubMed
-
- Blair HT, Sharp PE. Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat. Behav Neurosci. 1996;110:643–660. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources