

Uterine decidual response occurs in estrogen receptor-alpha-deficient mice
- PMID: 10342861
- PMCID: PMC4273314
- DOI: 10.1210/endo.140.6.6825
Uterine decidual response occurs in estrogen receptor-alpha-deficient mice
Abstract
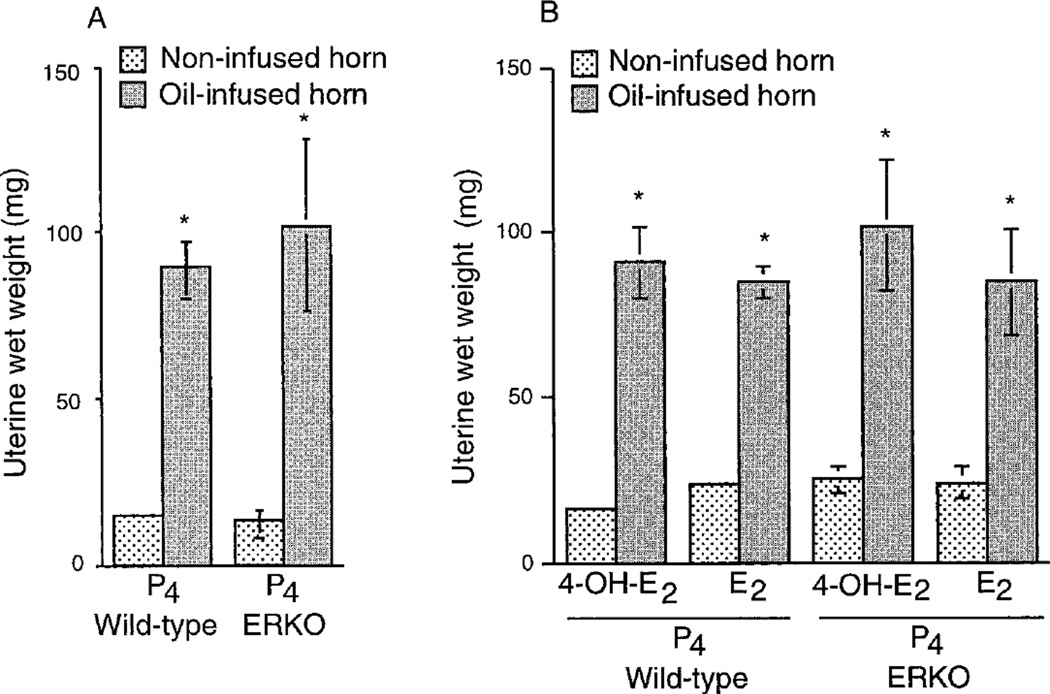
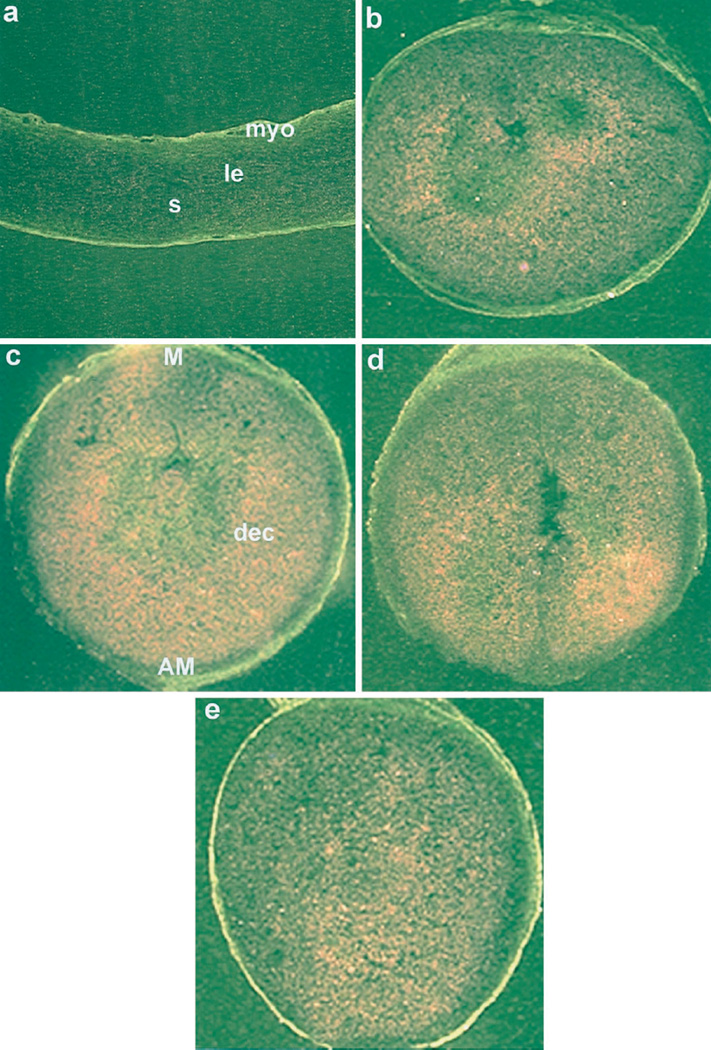
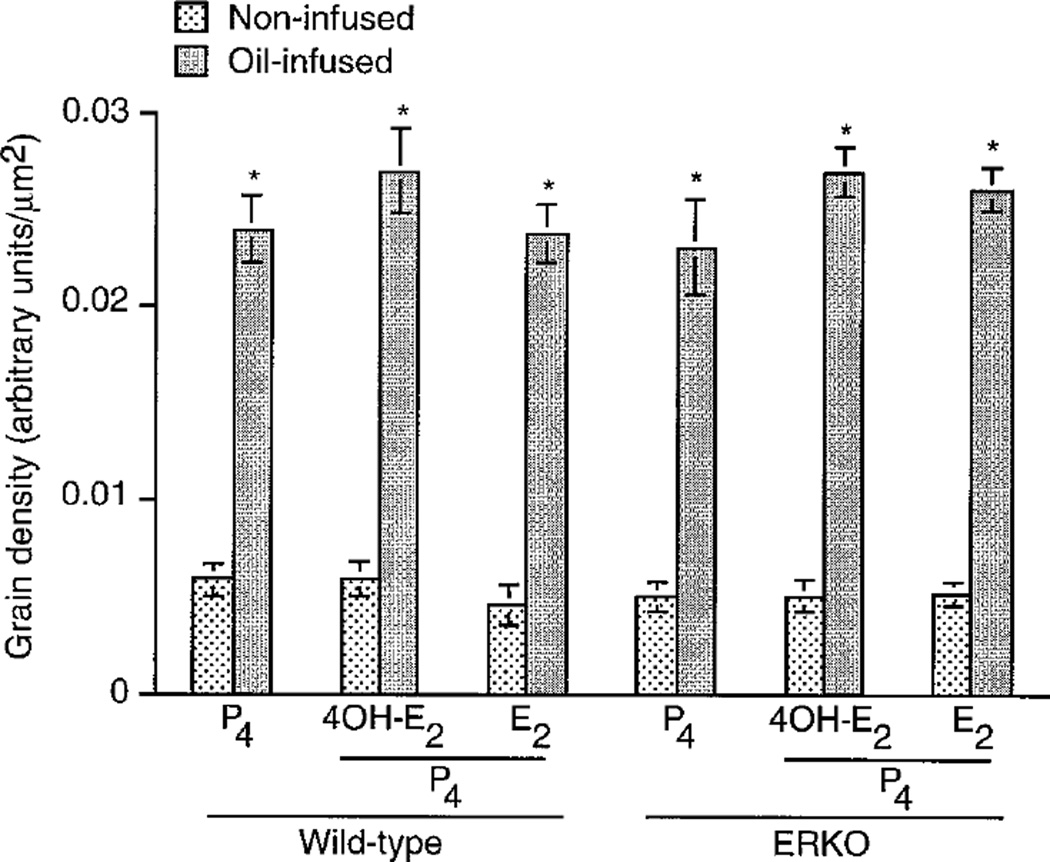
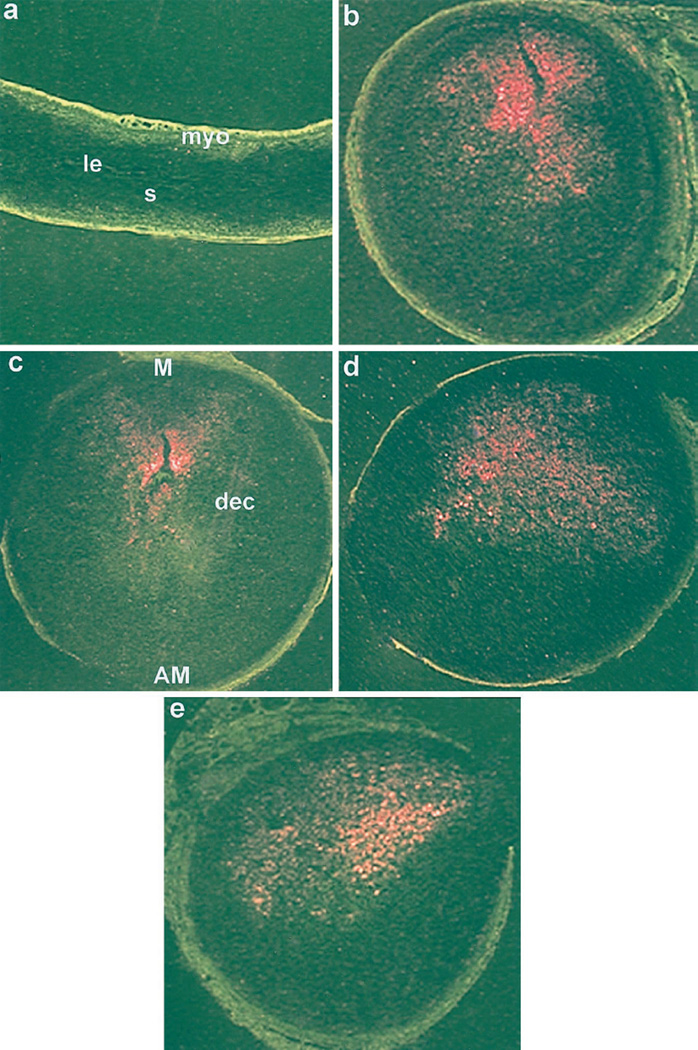
Embryo-uterine interactions leading to the attachment reaction is followed by stromal cell proliferation and differentiation into decidual cells (decidualization) at the sites of blastocyst apposition. In rodents, decidualization is also induced by application of an artificial stimulus (intraluminal oil infusion) in a pseudopregnant uterus, or to one that has been appropriately prepared by exogenous progesterone (P4) and estrogen. The process of decidualization is under the control of these steroids in the presence of blastocysts or deciduogenic stimuli. Although it is well known that estrogen is required for the induction of progesterone receptors in the uterus, the functional importance of estrogen in the process of decidualization is poorly understood. To better understand the role of estrogenic actions in decidualization, we used wild-type and estrogen receptor-alpha knock-out (ERKO) mice for induction of decidualization employing a defined steroid hormonal treatment schedule. Our results demonstrate that P4 alone induces decidualization in ovariectomized wild-type or ERKO mice in response to intraluminal oil infusion in the absence of estrogen. A combined treatment of either estradiol-17beta (E2) or its catecholmetabolite 4-hydroxyestradiol-17beta(4-OH-E2) with P4 does not potentiate the decidual response produced by P4 treatment alone in either ovariectomized wild-type or ERKO mice. The induction of decidual response was associated with up-regulation of decidual cell marker genes, such as progesterone receptor, metallothionein-1, and cyclooxygenase-2. The results suggest that the stromal cell sensitivity to decidualization is critically dependent on P4-regulated events, and estrogenic induction of progesterone receptor via classical nuclear ER-alpha is not critical for this process.
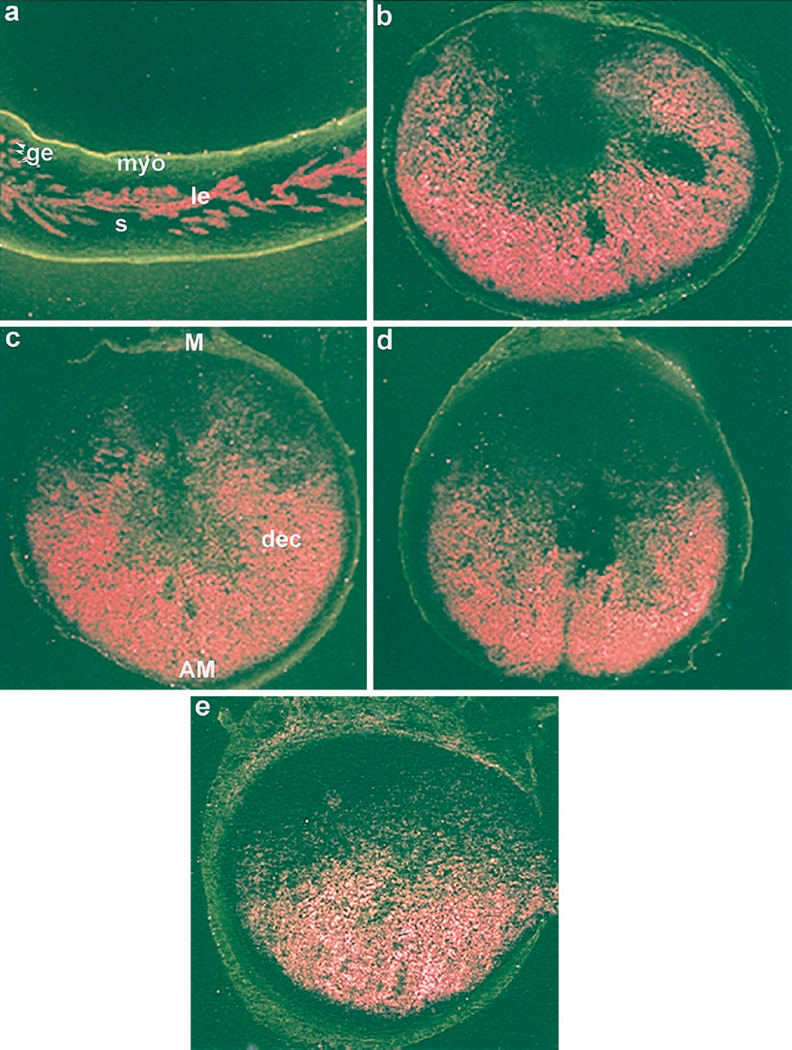
Figures
Similar articles
-
Studies using the estrogen receptor alpha knockout uterus demonstrate that implantation but not decidualization-associated signaling is estrogen dependent.Biol Reprod. 2002 Oct;67(4):1268-77. doi: 10.1095/biolreprod67.4.1268. Biol Reprod. 2002. PMID: 12297545
-
Hoxa-10 regulates uterine stromal cell responsiveness to progesterone during implantation and decidualization in the mouse.Mol Endocrinol. 1999 Jun;13(6):1005-17. doi: 10.1210/mend.13.6.0284. Mol Endocrinol. 1999. PMID: 10379898
-
Regulation of progesterone receptors and decidualization in uterine stroma of the estrogen receptor-alpha knockout mouse.Biol Reprod. 2001 Jan;64(1):272-83. doi: 10.1095/biolreprod64.1.272. Biol Reprod. 2001. PMID: 11133684
-
Implantation in the baboon: endometrial responses.Semin Reprod Endocrinol. 1999;17(3):257-65. doi: 10.1055/s-2007-1016233. Semin Reprod Endocrinol. 1999. PMID: 10797944 Review.
-
Progesterone action and responses in the alphaERKO mouse.Steroids. 2000 Oct-Nov;65(10-11):551-7. doi: 10.1016/s0039-128x(00)00113-6. Steroids. 2000. PMID: 11108859 Review.
Cited by
-
Physiological and molecular determinants of embryo implantation.Mol Aspects Med. 2013 Oct;34(5):939-80. doi: 10.1016/j.mam.2012.12.011. Epub 2013 Jan 2. Mol Aspects Med. 2013. PMID: 23290997 Free PMC article. Review.
-
Estrogen replacement regimen and brain infusion of lipopolysaccharide differentially alter steroid receptor expression in the uterus and hypothalamus.Endocrine. 2007 Dec;32(3):317-28. doi: 10.1007/s12020-008-9038-1. Epub 2008 Feb 5. Endocrine. 2007. PMID: 18247162
-
Uterine Patterning, Endometrial Gland Development, and Implantation Failure in Mice Exposed Neonatally to Genistein.Environ Health Perspect. 2020 Mar;128(3):37001. doi: 10.1289/EHP6336. Epub 2020 Mar 18. Environ Health Perspect. 2020. PMID: 32186404 Free PMC article.
-
Trophoblast differentiation during embryo implantation and formation of the maternal-fetal interface.J Clin Invest. 2004 Sep;114(6):744-54. doi: 10.1172/JCI22991. J Clin Invest. 2004. PMID: 15372095 Free PMC article. Review.
-
Differential uterine expression of estrogen and progesterone receptors correlates with uterine preparation for implantation and decidualization in the mouse.Endocrinology. 1999 Nov;140(11):5310-21. doi: 10.1210/endo.140.11.7148. Endocrinology. 1999. PMID: 10537162 Free PMC article.
References
-
- Psychoyos A. Endocrine control of egg implantation. In: Greep RO, Astwood EG, Geiger SR, editors. Handbook of Physiology. Washington, DC: American Physiological Society; 1973. pp. 187–215.
-
- Huet-Hudson YM, Andrews GK, Dey SK. Cell-type-specific localization of c-Myc protein in the mouse uterus: modulation by steroid hormones and analysis of the periimplantation period. Endocrinology. 1989;125:1683–1690. - PubMed
-
- Dey SK. Implantation. In: Adashi EY, Rock JA, Rosenwaks Z, editors. Reproductive Endocrinology, Surgery, and Technology. New York: Lippincott-Raven; 1996. pp. 421–434.
-
- De-Feo VJ. Decidualization. In: Wynn RM, editor. Cellular Biology of the uterus. New York: Appleton-Century-Crofts; 1967. pp. 191–205.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
