

doi: 10.1101/gad.13.10.1289.
Assembly of functionally active Drosophila origin recognition complex from recombinant proteins
Affiliations
- PMID: 10346817
- PMCID: PMC316721
- DOI: 10.1101/gad.13.10.1289
Item in Clipboard
Assembly of functionally active Drosophila origin recognition complex from recombinant proteins
Genes Dev.
.
Abstract
In eukaryotes the sites for the initiation of chromosomal DNA replication are believed to be determined in part by the binding of a heteromeric origin recognition complex (ORC) to DNA. We have cloned the genes encoding the subunits of the Drosophila ORC. Each of the genes is unique and can be mapped to discrete chromosomal locations implying that the pattern and developmental regulation of origin usage in Drosophila is not regulated solely by a large family of different ORC proteins. The six-subunit ORC can be reconstituted with recombinant proteins into a complex that restores DNA replication in ORC-depleted Drosophila or Xenopus egg extracts.
Figures
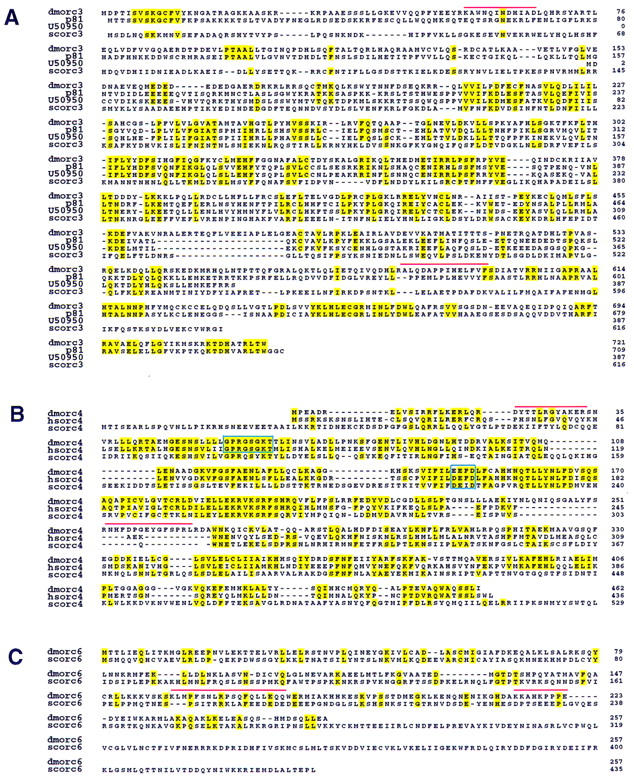
Drosophila homologs of S. cerevisiae, X. laevis, and H. sapiens ORC proteins. (A) The deduced amino acid sequence encoded by the DmORC3 is shown in alignment with S. cerevisiae ORC3, X. laevis p81, and an expressed sequence tag obtained from GenBank (accession no. U50950). Identical residues are highlighted. Red bars indicate tryptic polypeptides that were used for creation of the degenerate PCR primers used for cloning as described. (B) The deduced amino acid sequence encoded by DmORC4 is shown in alignment with S. cerevisiae ORC4 and H. sapiens ORC4. The putative nucleotide binding site and hydrolysis motifs are boxed. (C) The deduced amino acid sequence encoded by DmORC6 is shown in alignment with S. cerevisiae ORC6.
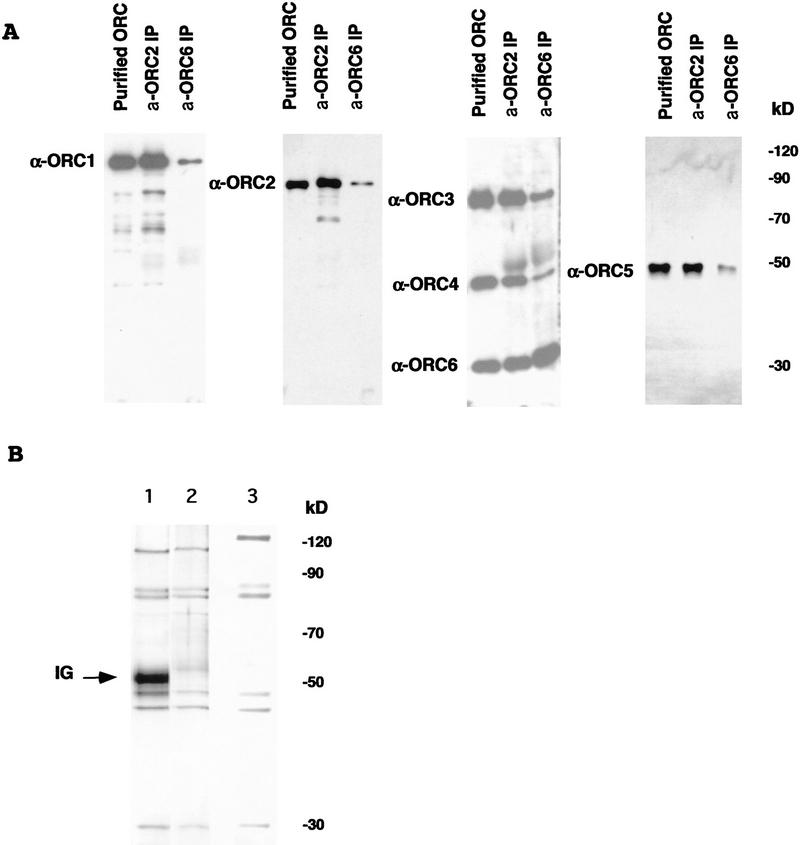
(A) Immunoblot analysis of six-subunit Drosophila ORC using antibodies raised against individual ORC subunits. Protein samples (∼20 ng of total ORC estimated by silver-stained material in the samples) corresponding to purified Drosophila embryonic ORC and immunoprecipitated by α-ORC2 or α-ORC6 antibodies, material was separated using SDS-PAGE and transferred to a PVDF membrane. Membrane strips were incubated with antibodies raised against individual Drosophila ORC subunits as indicated by the side labels. Signals with anti-ORC3 and anti-ORC6 sera were obtained first and the same filter was subsequently probed with the ORC4 reagent. Low levels of specific ORC1 and ORC2 proteolytic fragments are also visible. Protein bands were visualized by subsequent ECL assay. (B) Silver-stained gel of Drosophila ORC immunoprecipitated by α-ORC2 (lane 1) and recombinant baculovirus-expressed Drosophila ORC as eluted from a Ni column (lane 2) and subsequently purified on a glycerol gradient (lane 3). Lanes 1 and 2 are taken from the same gel. DmORC1 has an altered electophoretic mobility due to addition of a His tag.
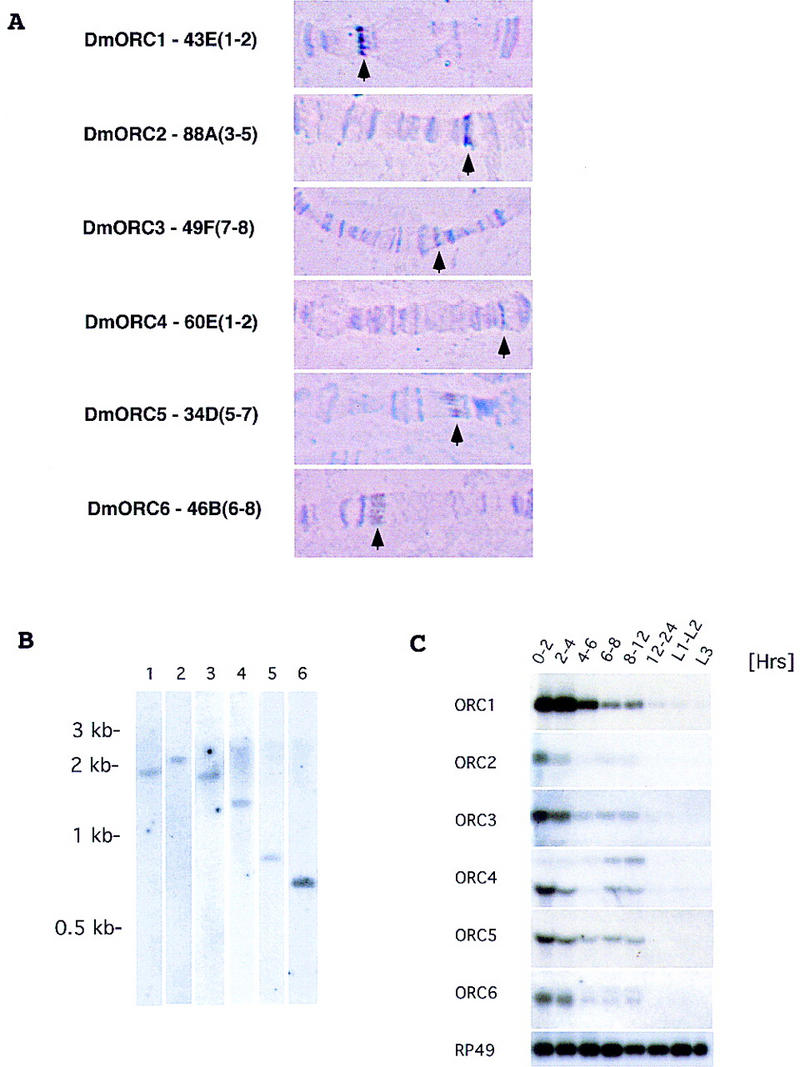
(A) Cytogenetic localization of Drosophila ORC genes based on results of fluorescent in situ hybridization. Chromosomal assignments for each gene are provided along with a representative picture of the polytene bands in the region of assignment (arrows point to specific bands). (B) Genomic blot using specific cDNA probes for Drosophila ORC genes. Genomic DNA (10 μg) was digested with HindIII and KpnI, separated on 0.8% agarose gel, and transferred to Zeta-probe blotting membrane (Bio-Rad). Membranes were hybridized with specific cDNA probes for each of the Drosophila ORC genes: (Lane 1) ORC1; (lane 2) ORC2; (lane 3) ORC3; (lane 4) ORC4; (lane 5) ORC5; (lane 6) ORC6. (C) Expression of Drosophila ORC genes during development. Northern blot analysis of total RNA isolated from Drosophila embryos and larvae extracted at different times of development was performed using radiolabeled probes corresponding to the individual ORC genes. RP49 (r ibosomal p rotein 49 ) serves as a loading control throughout. Sizes of mRNAs for individual ORC genes are as follows—ORC1, 2.7 kb; ORC2, 2.2 kb; ORC3, 2.3 kb; ORC4, 2.3 and 1.5 kb; ORC5, 1.5 kb; ORC6, 1.2 and 1.1 kb—and were estimated by comparison to a standard set of markers (GIBCO BRL RNA ladder).
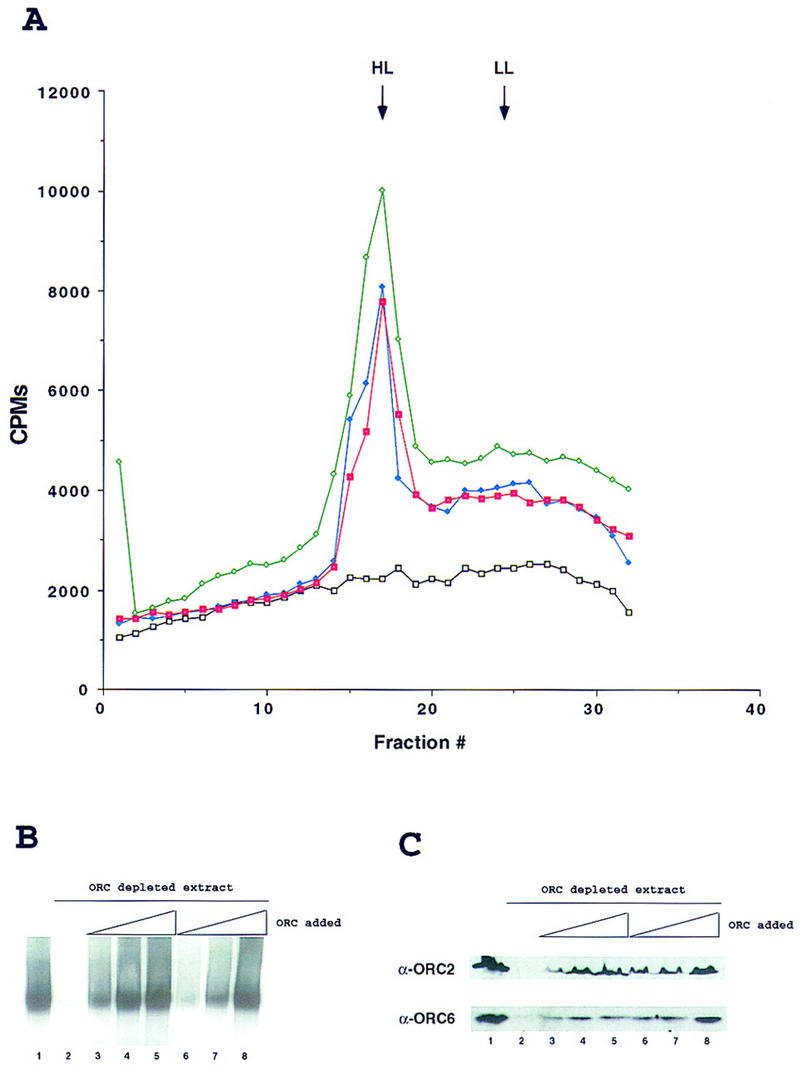
In vitro replication in Drosophila egg extracts is ORC dependent. (A) Density substitution analysis of replicated DNA. Demembraned Xenopus sperm DNA was incubated for 1 hr in Drosophila egg extract at a concentration of 10 ng/μl in the presence of BrdUTP and [α-32P]dCTP. DNA was extracted and subjected to centrifugation through a gradient of CsCl. Xenopus sperm DNA in Drosophila egg extract (green), in ORC-depleted Drosophila extract (black) and after addition to ORC-depleted extract of 100 ng of purified DmORC (blue) or baculovirus-expressed recombinant DmORC (red) are presented on the density profiles. (B) DNA replication in Drosophila extracts. Xenopus sperm DNA was incubated for 1 hr in Drosophila extract at a concentration of 2–5 ng/μl in the presence of [α-32P]dCTP. Where indicated, extracts were depleted for ORC using antibodies raised against DmORC2 and DmORC6. The reconstitution experiment was performed by addition of increasing amounts of purified DmORC [(lane 3) 25 ng; (lane 4) 50 ng; (lane 5) 100 ng] or recombinant baculovirus-expressed DmORC [(lane 6) 25 ng; (lane 7) 50 ng; (lane 8) 100 ng]. (C) Control for depletion and reconstitution experiments. Extracts used for in vitro replication in A and B were subjected to immunoblot analysis using antibodies raised against ORC2 and ORC6 (same order of lanes as in B).
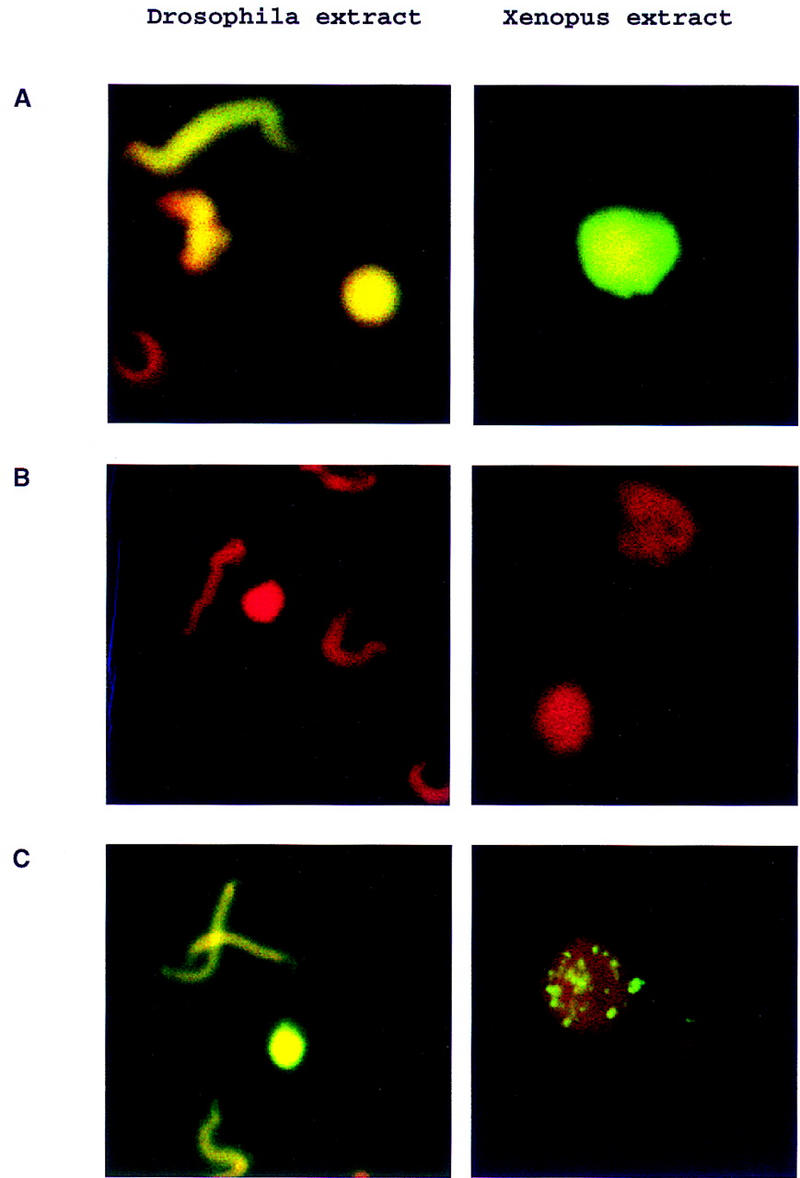
Visualization of ORC-dependent DNA replication in Drosophila and Xenopus extracts. Xenopus sperm DNA (10 ng/μl) was incubated for 2 hr in Drosophila or Xenopus egg extracts in the presence of biotin–16-UTP. In Drosophila, the extent of the sperm decondensation was variable and many of the replicating chromatin structures were elongated in shape but were clearly decondensed relative to sperm. The reactions were fixed and stained for detection of the incorporated UTP analog with fluorescein-conjugated streptavidin. DNA was counterstained with propidium iodide. Merged confocal images are presented. (A) DNA replication in mock-depleted Drosophila or Xenopus egg extracts; (B) DNA replication in ORC-depleted Drosophila or Xenopus egg extracts; (C) DNA replication in ORC-depleted Drosophila or Xenopus egg extract after readdition of purified DmORC (100 ng).
Similar articles
-
Interaction between the origin recognition complex and the replication licensing system in Xenopus.Cell. 1996 Oct 18;87(2):287-96. doi: 10.1016/s0092-8674(00)81346-x. Cell. 1996. PMID: 8861912
-
Role for a Xenopus Orc2-related protein in controlling DNA replication.Nature. 1996 Jan 25;379(6563):357-60. doi: 10.1038/379357a0. Nature. 1996. PMID: 8552193
-
Forced binding of the origin of replication complex to chromosomal sites in Drosophila S2 cells creates an origin of replication.J Cell Sci. 2012 Feb 15;125(Pt 4):965-72. doi: 10.1242/jcs.094409. Epub 2012 Mar 15. J Cell Sci. 2012. PMID: 22421364
-
Multiple functions of the origin recognition complex.Int Rev Cytol. 2007;256:69-109. doi: 10.1016/S0074-7696(07)56003-1. Int Rev Cytol. 2007. PMID: 17241905 Review.
-
ORC binding, gene amplification, and the nature of metazoan replication origins.Genes Dev. 1999 Oct 15;13(20):2619-23. doi: 10.1101/gad.13.20.2619. Genes Dev. 1999. PMID: 10541547 Review. No abstract available.
Cited by
-
Mechanisms and regulation of DNA replication initiation in eukaryotes.Crit Rev Biochem Mol Biol. 2017 Apr;52(2):107-144. doi: 10.1080/10409238.2016.1274717. Epub 2017 Jan 17. Crit Rev Biochem Mol Biol. 2017. PMID: 28094588 Free PMC article. Review.
-
Genetic interaction of an origin recognition complex subunit and the Polycomb group gene MEDEA during seed development.Plant Cell. 2004 Apr;16(4):1035-46. doi: 10.1105/tpc.019059. Epub 2004 Mar 12. Plant Cell. 2004. PMID: 15020747 Free PMC article.
-
Studies of the properties of human origin recognition complex and its Walker A motif mutants.Proc Natl Acad Sci U S A. 2005 Jan 4;102(1):69-74. doi: 10.1073/pnas.0408690102. Epub 2004 Dec 23. Proc Natl Acad Sci U S A. 2005. PMID: 15618391 Free PMC article.
-
DNA topology, not DNA sequence, is a critical determinant for Drosophila ORC-DNA binding.EMBO J. 2004 Feb 25;23(4):897-907. doi: 10.1038/sj.emboj.7600077. Epub 2004 Feb 5. EMBO J. 2004. PMID: 14765124 Free PMC article.
-
The origin recognition complex is dispensable for endoreplication in Drosophila.Proc Natl Acad Sci U S A. 2008 Aug 26;105(34):12343-8. doi: 10.1073/pnas.0805189105. Epub 2008 Aug 18. Proc Natl Acad Sci U S A. 2008. PMID: 18711130 Free PMC article.
References
-
- Aladjem MI, Rodewald LW, Kolman JL, Wahl GM. Genetic dissection of a mammalian replicator in the human beta-globin locus. Science. 1998;281:1005–1009. - PubMed
-
- Aparicio OM, Weinstein DM, Bell SP. Components and dynamics of DNA replication complexes in S. cerevisiae: Redistribution of MCM proteins and Cdc45p during S phase. Cell. 1997;91:59–69. - PubMed
-
- Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature. 1992;357:128–134. - PubMed
-
- Blow JJ, Laskey RA. Initiation of DNA replication in nuclei and purified DNA by a cell-free extract of Xenopus eggs. Cell. 1986;47:577–587. - PubMed
-
- ————— A role for the nuclear envelope in controling DNA replication within the cell cycle. Nature. 1988;332:546–548. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases