

IKK1-deficient mice exhibit abnormal development of skin and skeleton
- PMID: 10346820
- PMCID: PMC316728
- DOI: 10.1101/gad.13.10.1322
IKK1-deficient mice exhibit abnormal development of skin and skeleton
Abstract
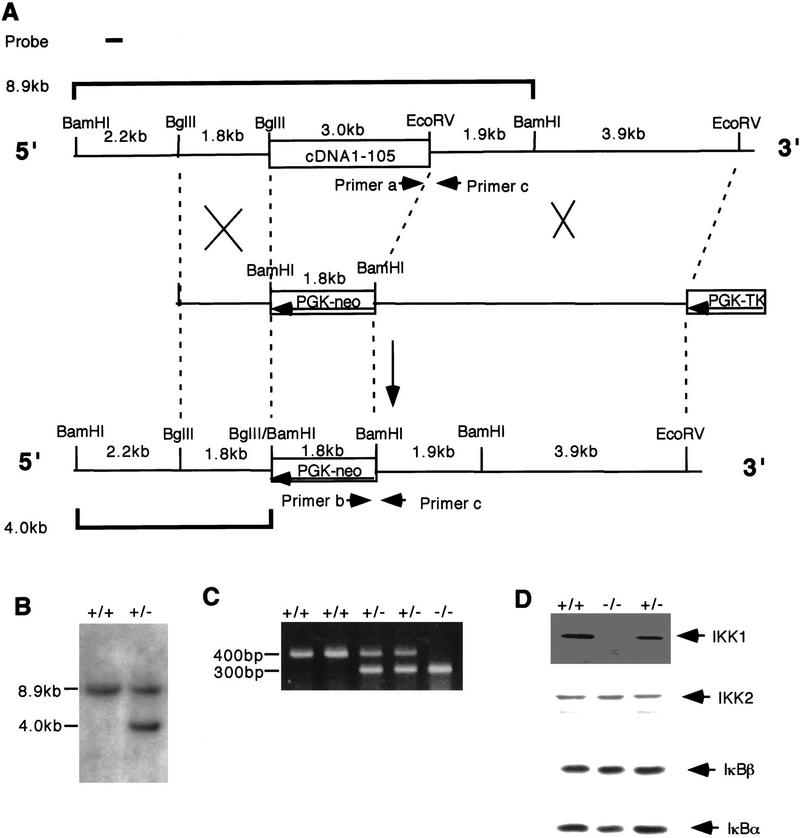
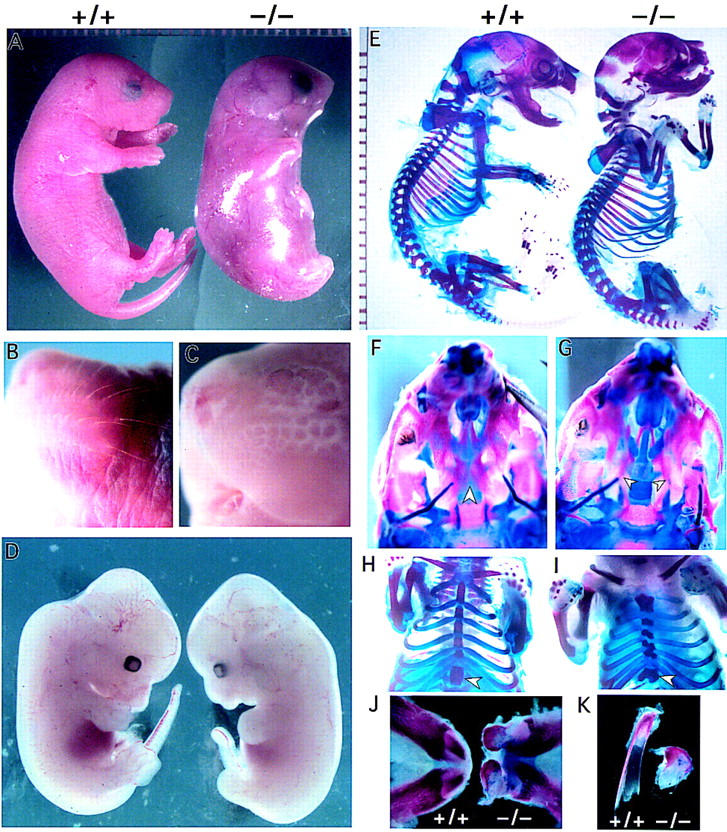
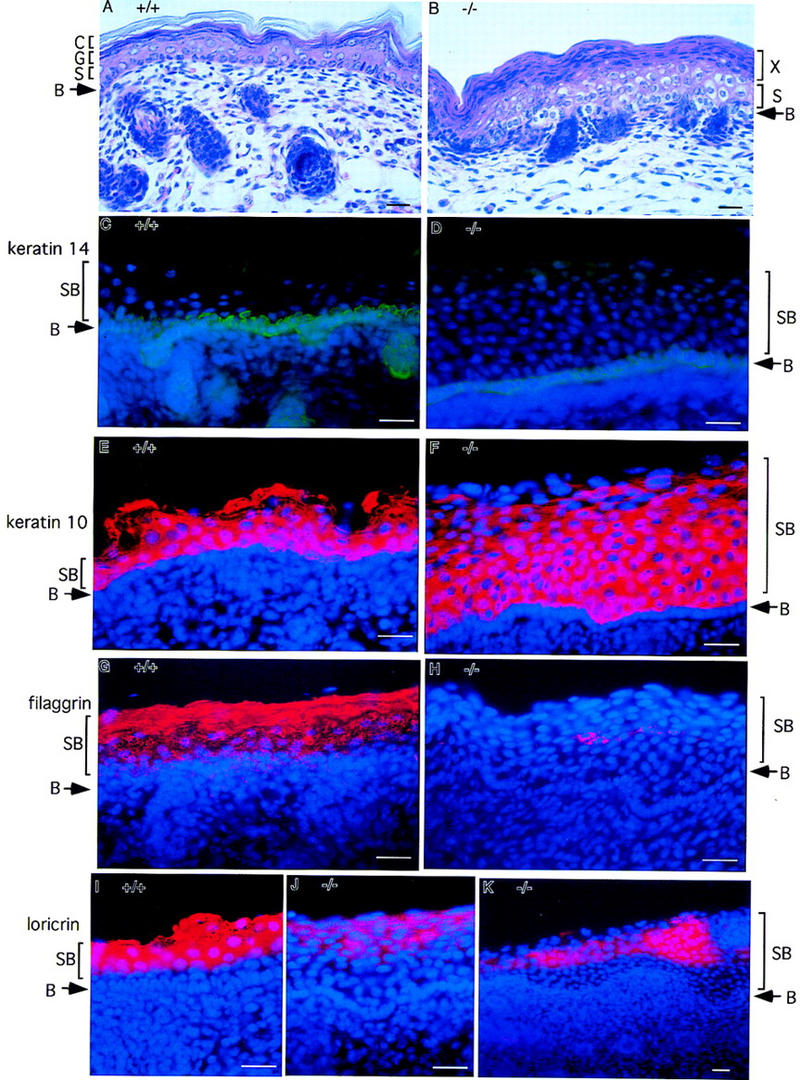
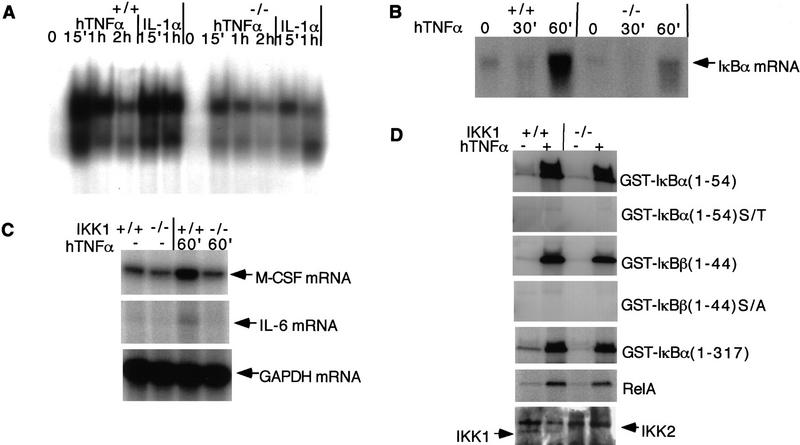
IkappaB kinases (IKKs) IKK1 and IKK2 are two putative IkappaBalpha kinases involved in NF-kappaB activation. To examine the in vivo functions of IKK1, we generated IKK1-deficient mice. The mutant mice are perinatally lethal and exhibit a wide range of developmental defects. Newborn mutant mice have shiny, taut, and sticky skin without whiskers. Histological analysis shows thicker epidermis, which is unable to differentiate. Limbs and tail are wrapped inside the skin and do not extend properly out of the body trunk. Skeleton staining reveals a cleft secondary palate, split sternebra 6, and deformed incisors. NF-kappaB activation mediated by TNFalpha and IL-1 is diminished in IKK1-deficient mouse embryonic fibroblast (MEF) cells. The IKK complex in the absence of IKK1 is capable of phosphorylating IkappaBalpha and IkappaBbeta in vitro. Our results support a role for IKK1 in NF-kappaB activation and uncover its involvement in skin and skeleton development. We conclude further that the two related kinases IKK1 and IKK2 have distinct functions and can not be substituted for each other's functions.
Figures
References
-
- Attar RM, Caamano J, Carrasco D, Iotsova V, Ishikawa H, Ryseck RP, Weih F, Bravo R. Genetic approaches to study Rel/NF-κB/I κB function in mice. Semin Cancer Biol. 1997;8:93–101. - PubMed
-
- Baeuerle PA, Baichwal VR. NF-κ B as a frequent target for immunosuppressive and anti-inflammatory molecules. Adv Immunol. 1997;65:111–137. - PubMed
-
- Beg AA, Baltimore D. An essential role for NF-κ in preventing TNF-alpha-induced cell death. Science. 1996;274:782–784. - PubMed
-
- Beg AA, Sha WC, Bronson RT, Ghosh S, Baltimore D. Embryonic lethality and liver degeneration in mice lacking the RelA component of NF-κ B. Nature. 1995;376:167–170. - PubMed
-
- Bushdid PB, Brantley DM, Yull FE, Blaeuer GL, Hoffman LH, Niswander L, Kerr LD. Inhibition of NF-κB activity results in disruption of the apical ectodermal ridge and aberrant limb morphogenesis. Nature. 1998;392:615–618. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases