

Mice devoid of all known thyroid hormone receptors are viable but exhibit disorders of the pituitary-thyroid axis, growth, and bone maturation
- PMID: 10346821
- PMCID: PMC316730
- DOI: 10.1101/gad.13.10.1329
Mice devoid of all known thyroid hormone receptors are viable but exhibit disorders of the pituitary-thyroid axis, growth, and bone maturation
Abstract
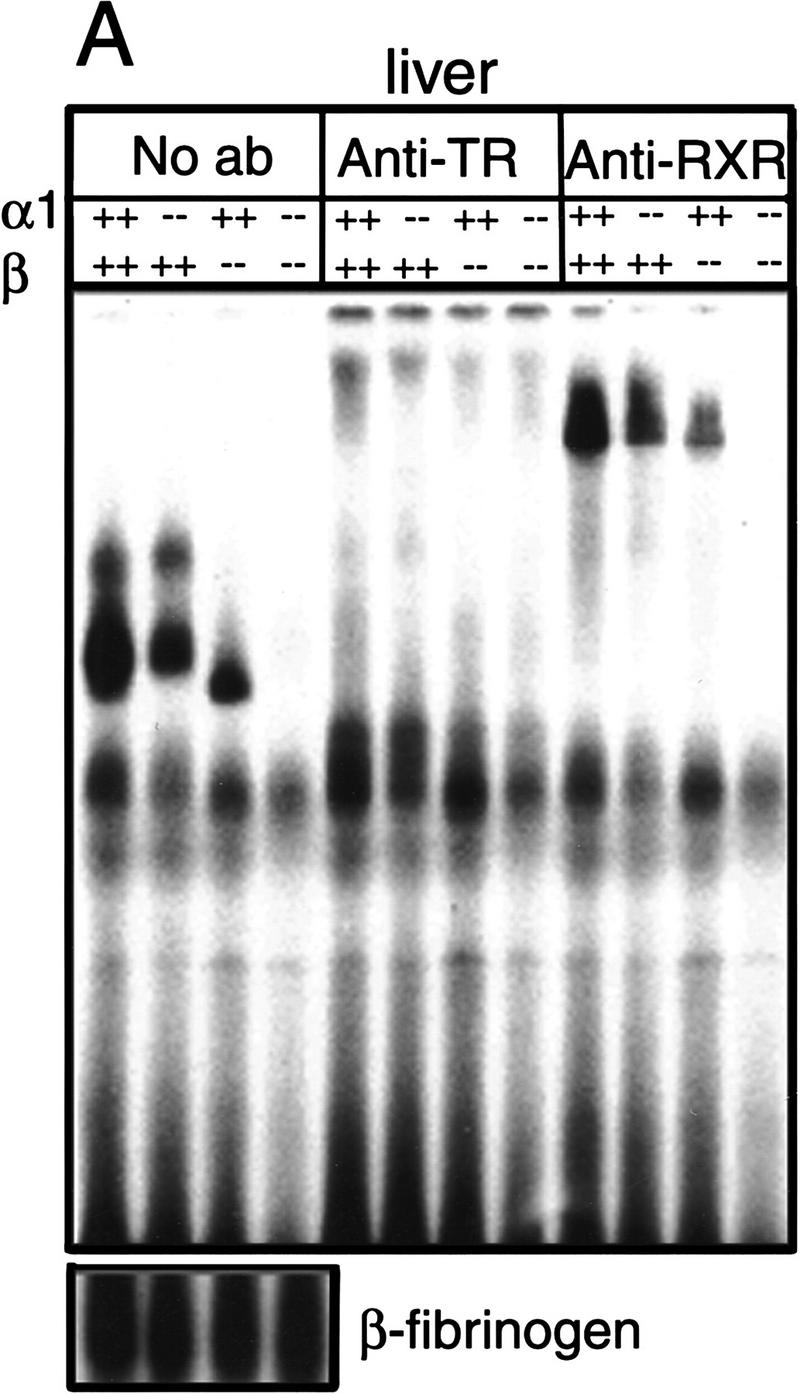
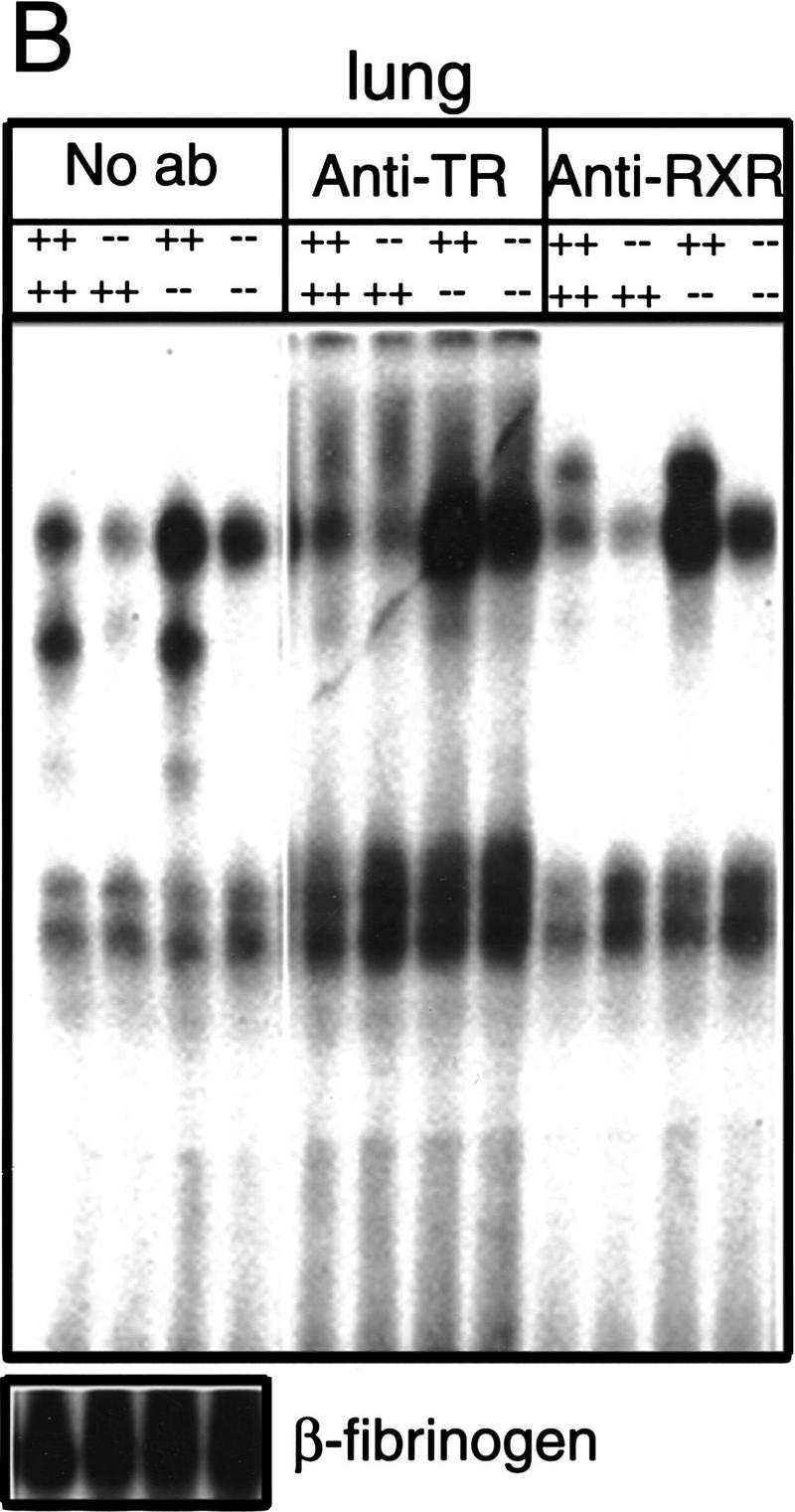
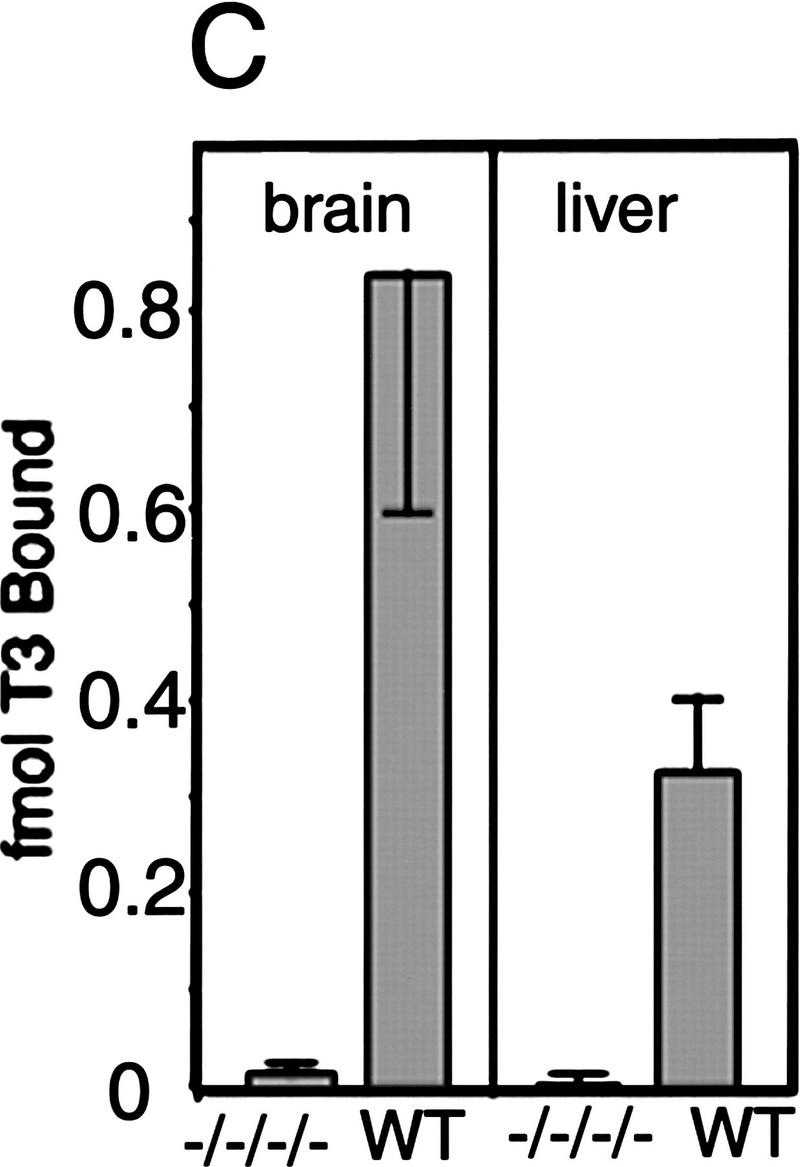
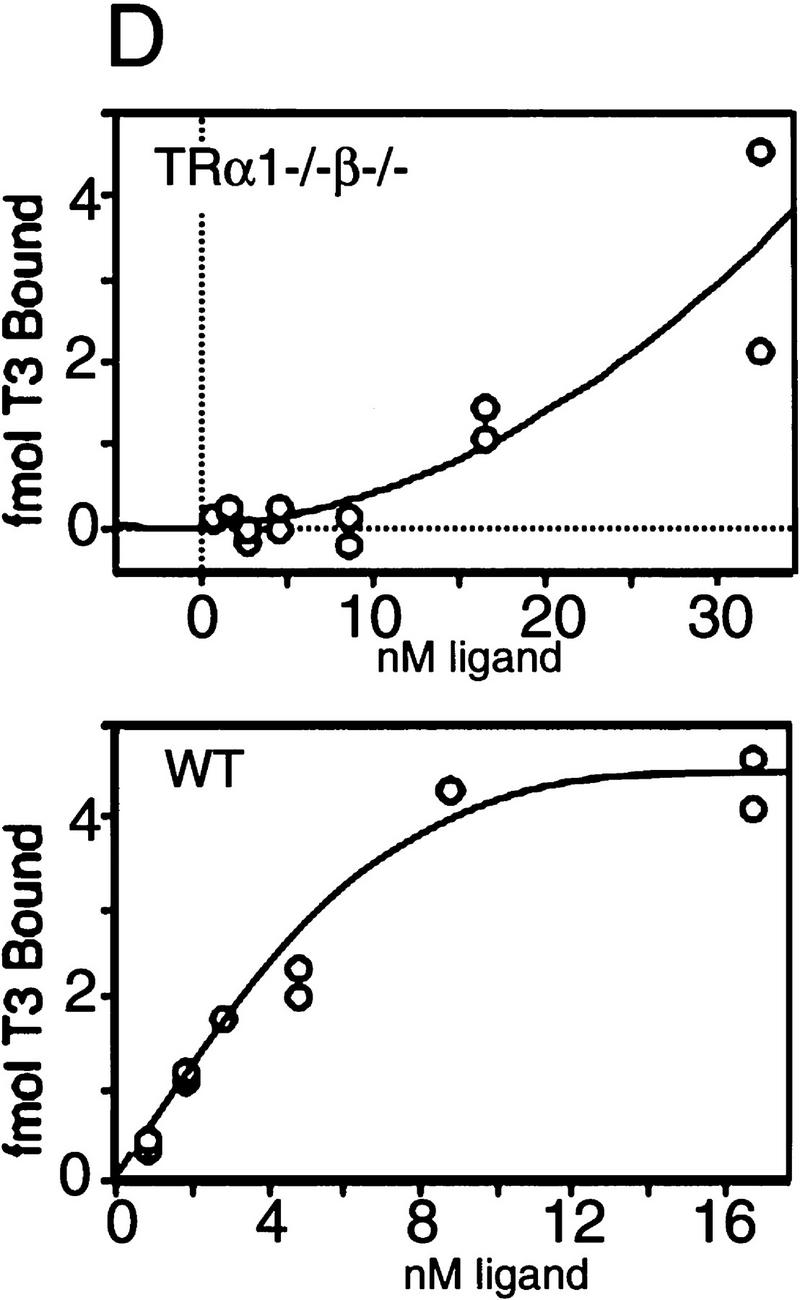
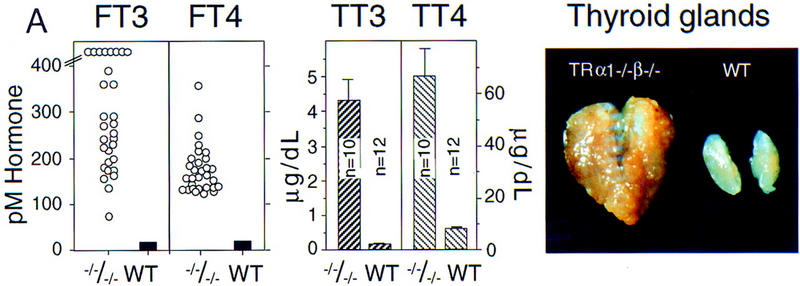
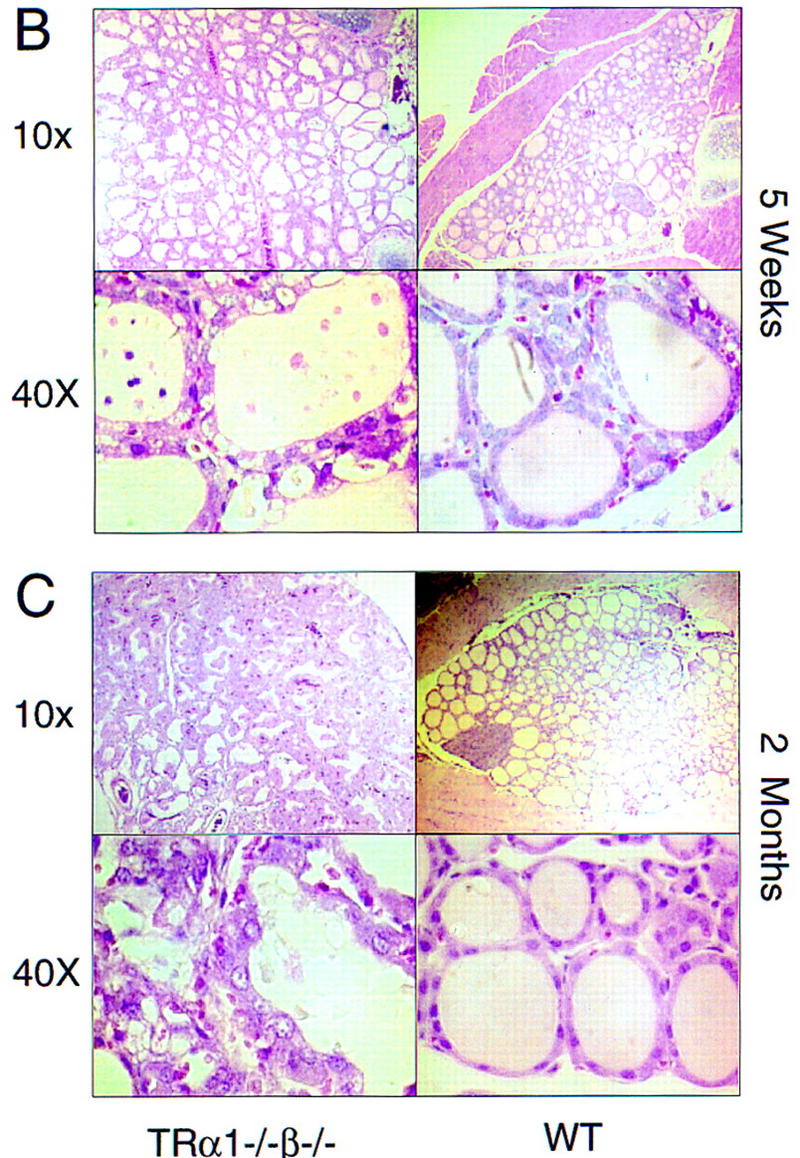
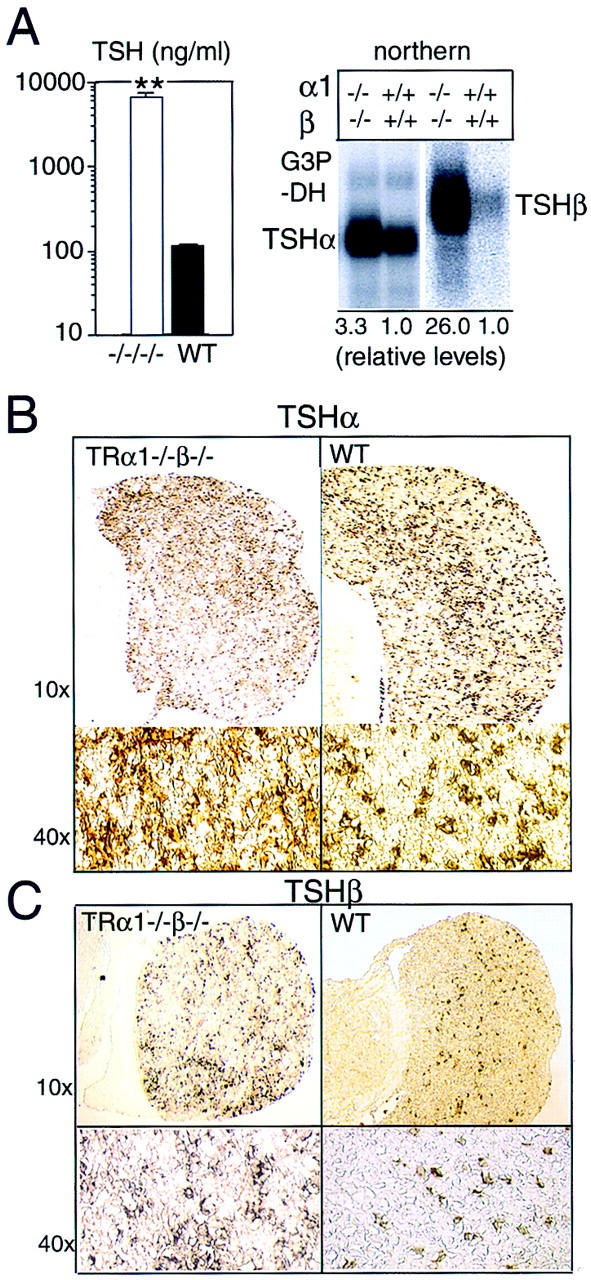
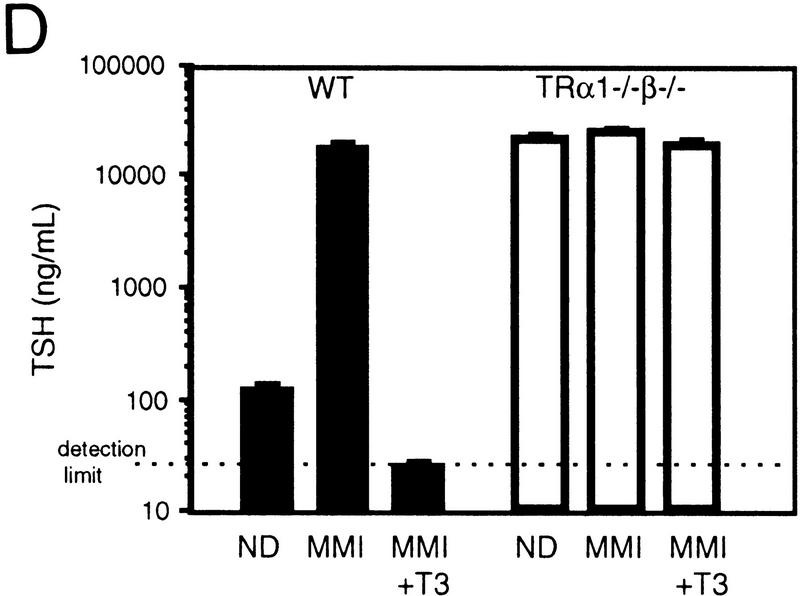
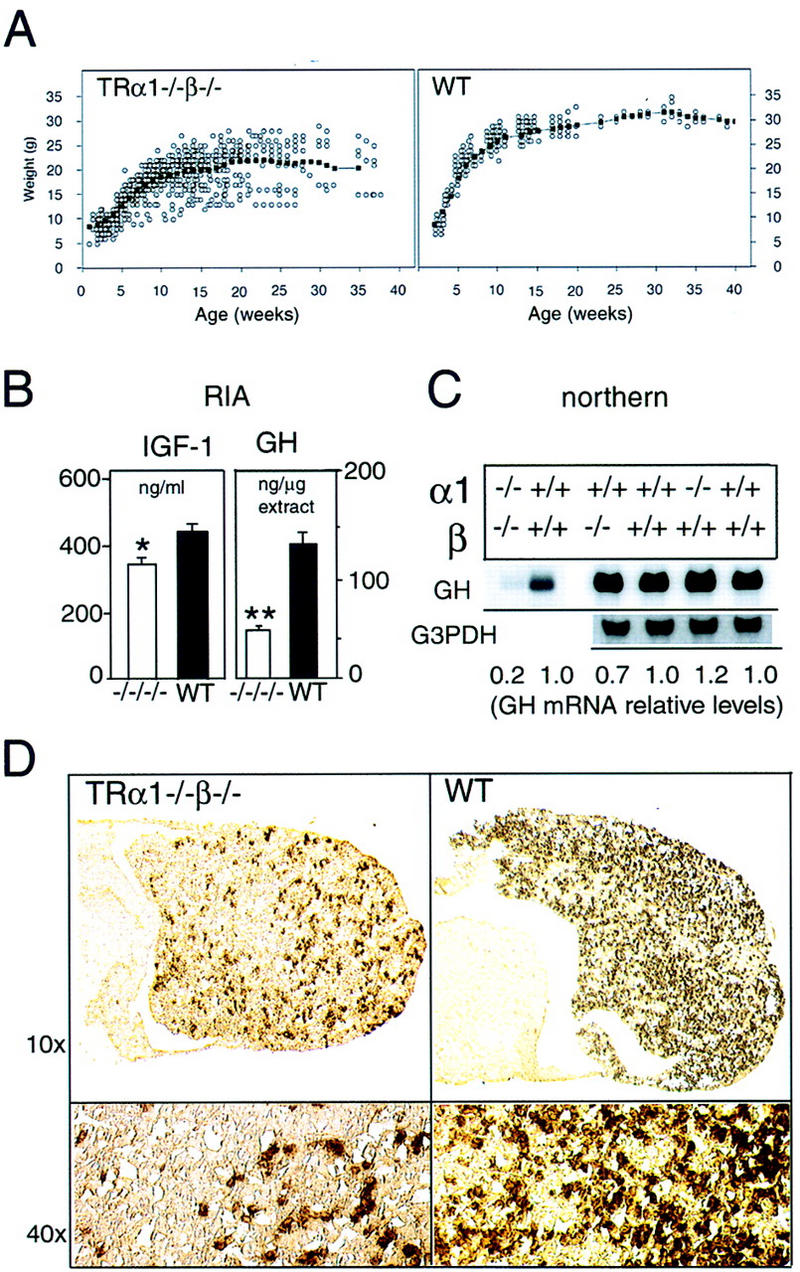
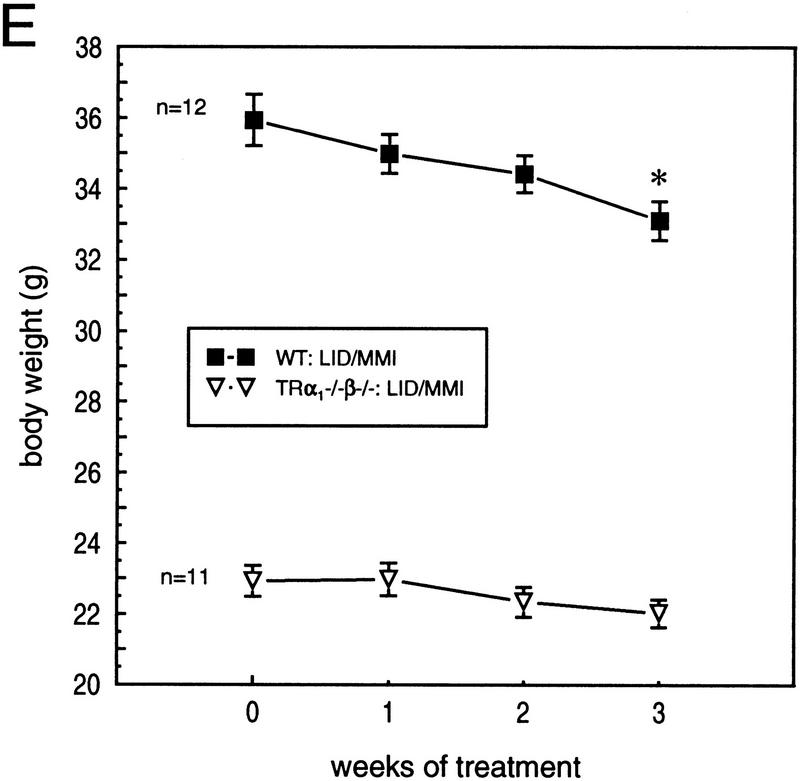
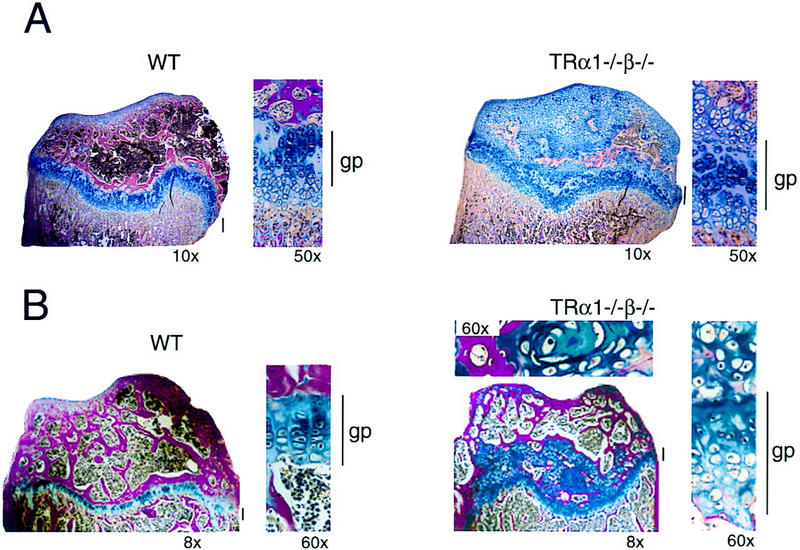
Thyroid hormone (T3) has widespread functions in development and homeostasis, although the receptor pathways by which this diversity arises are unclear. Deletion of the T3 receptors TRalpha1 or TRbeta individually reveals only a small proportion of the phenotypes that arise in hypothyroidism, implying that additional pathways must exist. Here, we demonstrate that mice lacking both TRalpha1 and TRbeta (TRalpha1(-/-)beta-/-) display a novel array of phenotypes not found in single receptor-deficient mice, including an extremely hyperactive pituitary-thyroid axis, poor female fertility and retarded growth and bone maturation. These results establish that major T3 actions are mediated by common pathways in which TRalpha1 and TRbeta cooperate with or substitute for each other. Thus, varying the balance of use of TRalpha1 and TRbeta individually or in combination facilitates control of an extended spectrum of T3 actions. There was no evidence for any previously unidentified T3 receptors in TRalpha1(-/-)beta-/- mouse tissues. Compared to the debilitating symptoms of severe hypothyroidism, the milder overall phenotype of TRalpha1(-/-)beta-/- mice, lacking all known T3 receptors, indicates divergent consequences for hormone versus receptor deficiency. These distinctions suggest that T3-independent actions of T3 receptors, demonstrated previously in vitro, may be a significant function in vivo.
Figures
Similar articles
-
Distinct tissue-specific roles for thyroid hormone receptors beta and alpha1 in regulation of type 1 deiodinase expression.Mol Endocrinol. 2001 Mar;15(3):467-75. doi: 10.1210/mend.15.3.0605. Mol Endocrinol. 2001. PMID: 11222747
-
Contrasting skeletal phenotypes in mice with an identical mutation targeted to thyroid hormone receptor alpha1 or beta.Mol Endocrinol. 2005 Dec;19(12):3045-59. doi: 10.1210/me.2005-0224. Epub 2005 Jul 28. Mol Endocrinol. 2005. PMID: 16051666
-
Requirement for thyroid hormone receptor beta in T3 regulation of cholesterol metabolism in mice.Mol Endocrinol. 2002 Aug;16(8):1767-77. doi: 10.1210/me.2002-0009. Mol Endocrinol. 2002. PMID: 12145333
-
Insight into the physiological actions of thyroid hormone receptors from genetically modified mice.J Endocrinol. 2002 Dec;175(3):553-70. doi: 10.1677/joe.0.1750553. J Endocrinol. 2002. PMID: 12475367 Review.
-
Isoform-dependent actions of thyroid hormone nuclear receptors: lessons from knockin mutant mice.Steroids. 2005 May-Jun;70(5-7):450-4. doi: 10.1016/j.steroids.2005.02.003. Epub 2005 Mar 16. Steroids. 2005. PMID: 15862829 Review.
Cited by
-
The immediate and late effects of thyroid hormone (triiodothyronine) on murine coagulation gene transcription.PLoS One. 2015 May 26;10(5):e0127469. doi: 10.1371/journal.pone.0127469. eCollection 2015. PLoS One. 2015. PMID: 26011296 Free PMC article.
-
Delayed development of specific thyroid hormone-regulated events in transthyretin null mice.Am J Physiol Endocrinol Metab. 2013 Jan 1;304(1):E23-31. doi: 10.1152/ajpendo.00216.2012. Epub 2012 Oct 23. Am J Physiol Endocrinol Metab. 2013. PMID: 23092911 Free PMC article.
-
Comparative Analysis of Transcriptome Profiles Reveals Distinct and Organ-Dependent Genomic and Nongenomic Actions of Thyroid Hormone in Xenopus tropicalis Tadpoles.Thyroid. 2023 Apr;33(4):511-522. doi: 10.1089/thy.2022.0469. Epub 2023 Jan 24. Thyroid. 2023. PMID: 36503276 Free PMC article.
-
Effects of ligand and thyroid hormone receptor isoforms on hepatic gene expression profiles of thyroid hormone receptor knockout mice.EMBO Rep. 2003 Jun;4(6):581-7. doi: 10.1038/sj.embor.embor862. EMBO Rep. 2003. PMID: 12776178 Free PMC article.
-
An unliganded thyroid hormone beta receptor activates the cyclin D1/cyclin-dependent kinase/retinoblastoma/E2F pathway and induces pituitary tumorigenesis.Mol Cell Biol. 2005 Jan;25(1):124-35. doi: 10.1128/MCB.25.1.124-135.2005. Mol Cell Biol. 2005. PMID: 15601836 Free PMC article.
References
-
- Aguanno A, Afar R, Albert V. Tissue-specific expression of the non-neuronal promoter of the aromatic l-amino acid decarboxylase gene is regulated by hepatocyte nuclear factor 1. J Biol Chem. 1996;271:4528–4538. - PubMed
-
- Beamer W, Eicher E, Maltais L, Southard J. Inherited primary hypothyroidism in mice. Science. 1981;212:61–63. - PubMed
-
- Beamer W, Maltais L, DeBaets M, Eicher E. Inherited congenital goiter in mice. Endocrinology. 1987;120:838–840. - PubMed
-
- Brent G, Moore D, Larsen P. Thyroid hormone regulation of gene expression. Annu Rev Physiol. 1991;53:17–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases