

doi: 10.1101/gad.13.10.1342.
Polarity determination in the Drosophila eye: a novel role for unpaired and JAK/STAT signaling
Affiliations
- PMID: 10346822
- PMCID: PMC316719
- DOI: 10.1101/gad.13.10.1342
Item in Clipboard
Polarity determination in the Drosophila eye: a novel role for unpaired and JAK/STAT signaling
Genes Dev.
.
Abstract
The JAK/STAT signaling pathway is required for many processes including cytokine signaling, hematopoiesis, gliagenesis, and Drosophila segmentation. In this report we present evidence demonstrating that the JAK/STAT pathway is also central to the establishment of planar polarity during Drosophila eye development. We show that a localized source of the pathway ligand, Unpaired, is present at the midline of the developing eye, which is capable of activating the JAK/STAT pathway over long distances. A gradient of JAK/STAT activity across the DV axis of the eye regulates ommatidial polarity via an unidentified second signal. Additionally, localized Unpaired influences the position of the equator via repression of mirror.
Figures
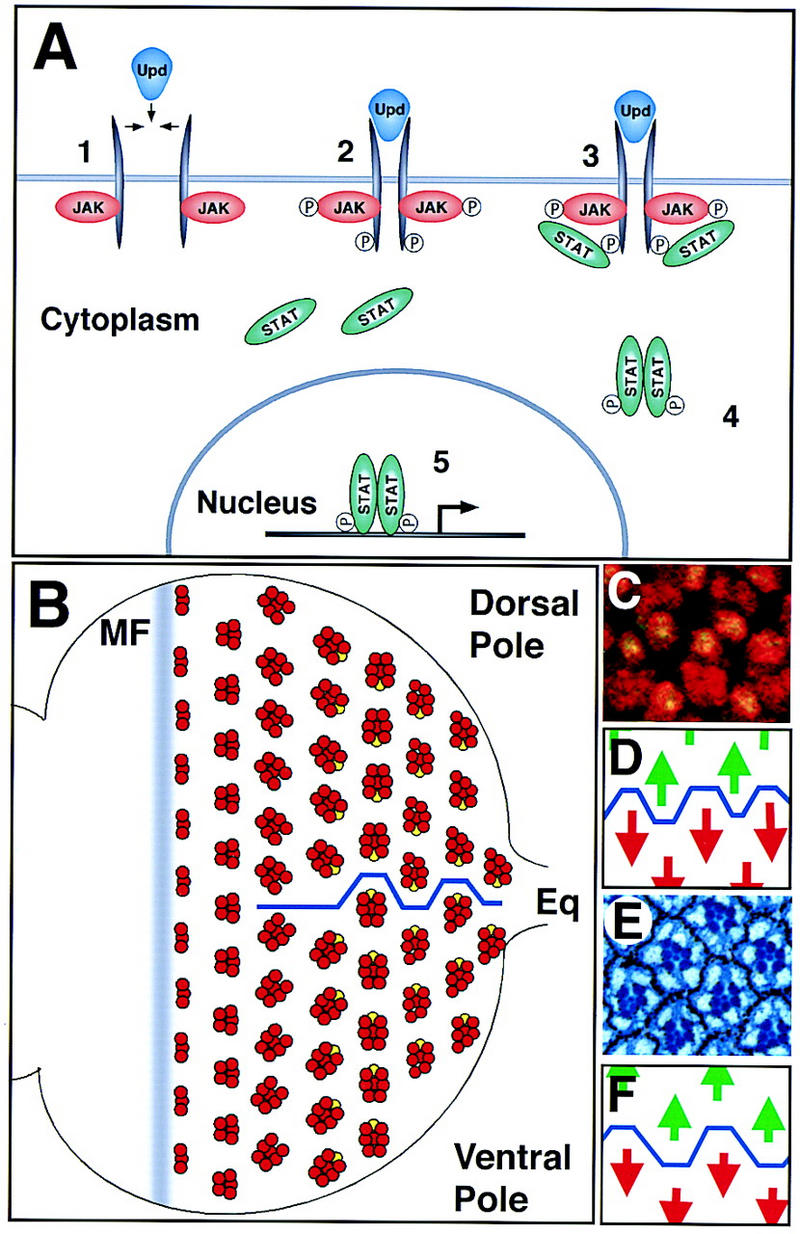
The JAK/STAT pathway and the establishment of dorsoventral polarity in the eye disc. Anterior is to the left and dorsal up, here and in all subsequent figures. (A) The transduction of extracellular signals from the cell membrane to the nucleus by the JAK/STAT signaling pathway. (B) A schematized representation of ommatidial development and rotation. The movement of the morphogenetic furrow (MF) from posterior to anterior across the disc leads to the determination of successive rows of increasingly mature ommatidial clusters (red). Shortly before determination of the R7 photoreceptor (yellow), dorsal and ventral clusters rotate in opposite directions to generate the equator (blue line). (C) Ommatidial clusters at the equator in the third instar eye disc. All photoreceptor nuclei are marked by anti-Elav (red) and the R7 nucleus is also marked by expression of Prospero (green, yellow when overlaid). (D) Schematic representation of C showing the position of the equator (blue line) through this region and the green dorsal arrows and red ventral arrows used to indicate ommatidial polarity. (E) Adult eye section at the equator showing the stereotypical trapezoidal arrangement of rhabdomeres in each adult ommatidial cluster. Each ommatidium is surrounded by a hexagonal array of pigment granules. (F) Schematic representation of E showing position of the equator (blue line) through this region and the green dorsal arrows and red ventral arrows used to indicate ommatidial polarity.
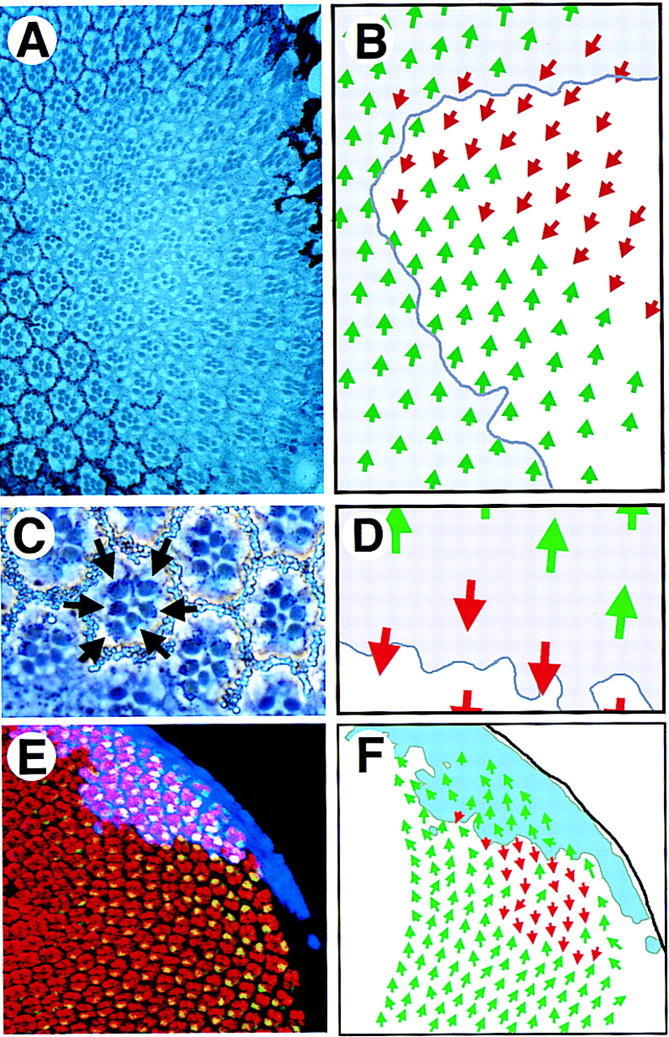
LOF and GOF JAK/STAT pathway clones show abnormal ommatidial polarity. (A) A large dorsal hop2 clone (marked by absence of white+ pigment granules) contains normally differentiated ommatidia in both correct and inverted orientation with respect to surrounding wild-type clusters. (B) Schematic representation of the clone shown in A. Dorsal-oriented ommatidia are shown as green arrows and inverted ommatidia are shown in red. The approximate position of the clonal boundary is shown by the purple line. A field of inverted ommatidia is present around the polar margin of the clone. (C) A high-magnification view of ommatidia along the polar boundary of a hop2 clone (marked by absence of white+) showing an inverted ommatidium comprised of entirely wild-type photoreceptor cells (note dark pigment granules associated with the rhabdomeres of wild-type photoreceptors, indicated by arrows). (D) Schematic representation of clone shown in C. Dorsal ommatidia are shown by green and ventral by red arrows. Boundary of the clone is indicated. (E) A clone of cells ectopically expressing Upd (blue) in the dorsal part of an eye disc stained to show Elav (red) and Prospero (green, yellow in overlay). Ommatidia with inverted orientation are seen on the ventral edge of the clone. (F) A schematic representation of clone shown in E indicating the extent of the Upd expressing clone (pale blue) and the ommatidial orientation indicated by dorsal green and ventral red arrows.
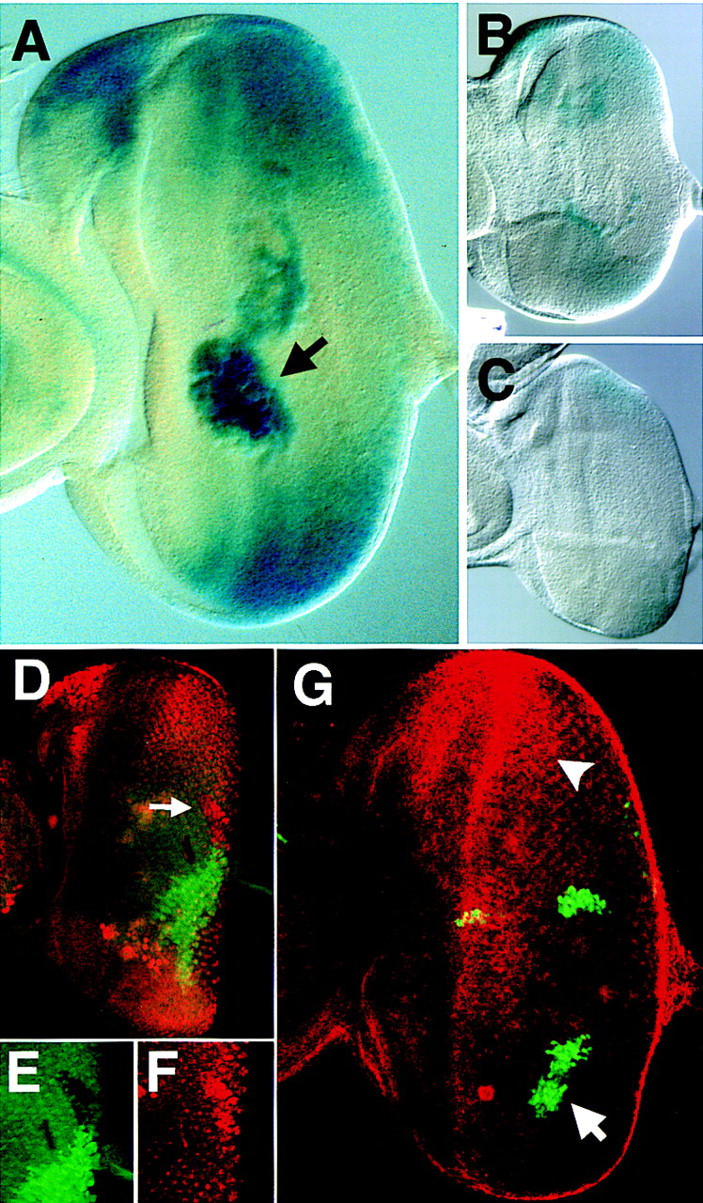
The expression of stat92E–lacZ is negatively regulated by the activity of JAK/STAT signaling. (A) stat92E–lacZ staining in a third instar disc showing highest levels of lacZ at the poles of the eye field, at the ocellar spot, and in macrophage cells located in the middle of the eye disc below the plane of focus (arrow). (B,C) Eye discs prepared and stained under identical conditions carrying stat92E–lacZ (B) and stat92E–lacZ in a hopTuml mutant background (C). (D–F) Double label of eye imaginal disc stained for stat92E–lacZ (red) and a GFP clonal marker (green). A hop2 clone marked by lack of green staining (arrow in D, enlarged in E) and results in higher levels of stat92E–lacZ expression autonomously within the clone (D, enlarged in F). (G) Upd and GFP (green) are coexpressed in GOF clones within the eye imaginal disc also stained to show the pattern of stat92E–lacZ (red). An Upd-expressing clone in the ventral portion of the disc (arrow) completely abolishes the stat92E–lacZ gradient in the adjacent tissue such that no staining is seen at the ventral pole. Staining at the dorsal pole remains strong (arrowhead). Large cells staining in the middle of the disc are macrophages.
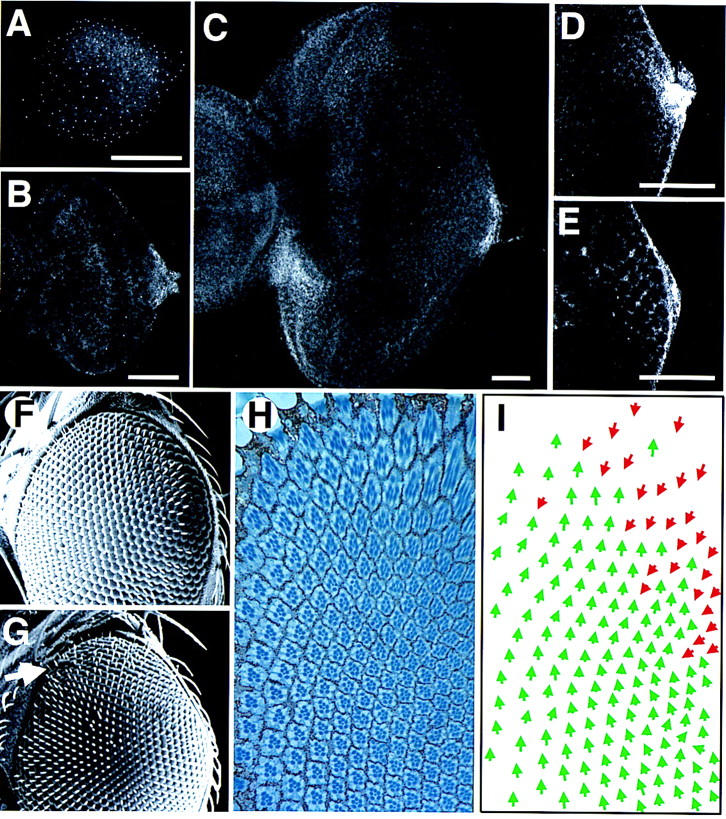
The wild-type expression pattern of Upd and the phenotype induced by ectopic expression of Upd from the poles of the eye. (A–E) The pattern of staining with an anti-Upd polyclonal antibody; scale bars, 50 μm. (A) First instar disc. Staining is faintly visible in a horseshoe around the poles and posterior margin of the disc. The small bright spots of staining distributed across the whole disc may represent staining in the peripodial membrane and/or could be nonspecific. (B) Second instar. Expression is localized to the posterior margin at the DV midline, adjacent to the optic stalk. (C) Late third instar. The furrow has progressed more than halfway across the disc. Expression is still seen localized adjacent to the optic stalk, however, a new region of high-level expression is evident on the ventro–anterior margin adjacent to the antennal disc, which is not present in younger discs (and thus presumably plays no role in ommatidial polarity determination). (D) Higher magnification view of the posterior midline region of an early third instar disc, focused basally. A small cluster of cells shows high-level cytoplasmic expression. (E) More apical view of disc shown in D. A gradient of extracellular Upd can be seen in the extracellular space between the nascent ommatidial clusters. (F) Scanning electron micrograph (SEM) of the dorsal half of a wild-type adult eye. (G) SEM of the dorsal half of an eye in which Upd expression was driven at the dorsal and ventral poles during development by the 30A–GAL4 driver at 18°C. The eye appears normal apart from a weak disruption of externally visible facet packing at the dorsal pole (arrow). (H) Section through the dorsal region of an eye similar to G produced by expression of Upd with the 30A–GAL4 driver. The eye is normal in the ventral part of the field (close to the equator), but inverted ommatidia are seen near the dorsal pole. (I) Schematic representation of C. Dorsal ommatidia are shown by green and ventral by red arrows. Note that there is a region of inversion of ommatidial polarity near the dorsal pole, but that ommatidia of both fates are clearly mixed.
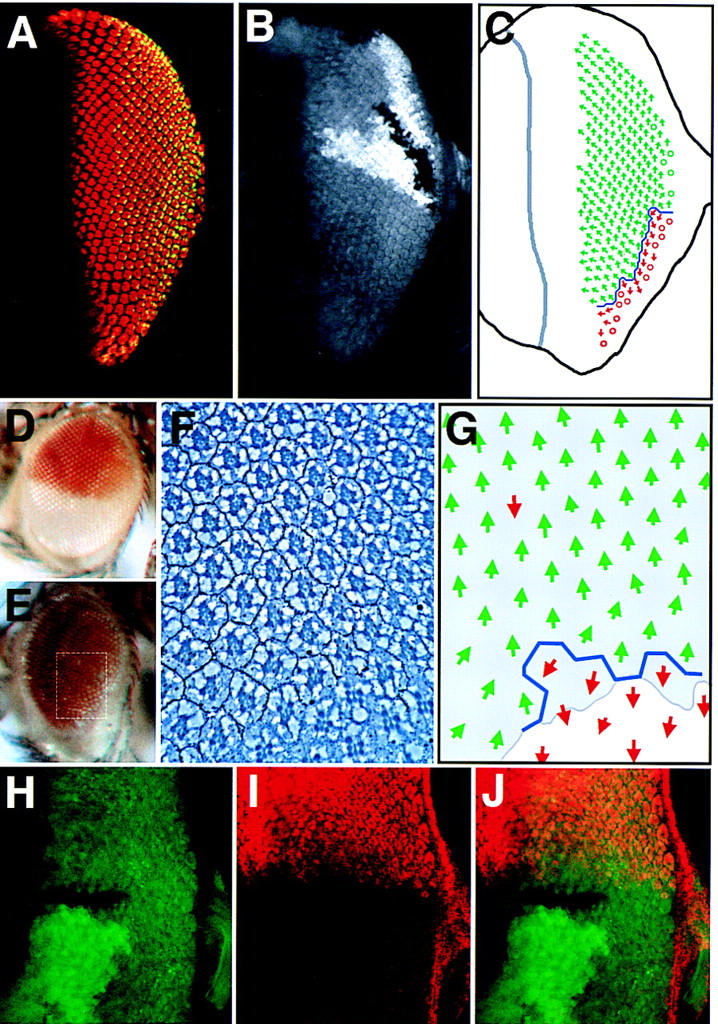
Upd is required for dorsal restriction of mirr expression and positioning of the equator, but hop clones do not affect mirr expression. (A,B) An eye disc containing an Upd clone situated at the posterior of the eye field is dorsalized with the equator shifted to the ventral margin. (A) The photoreceptor markers Elav (red) and Prospero (green, yellow in overlay); (B) The clonal marker in which nonstaining cells lack upd function. (C) Schematic representation of the disc shown in A and B. Much of the eye field has assumed the dorsal (green arrow) fate and only a small number of ventral ommatidia (red arrows) are present along the ventral posterior margin of the eye separated by an equator (blue line) (○). Ommatidia with unscoreable rotation. (D) A wild-type eye showing the dorsal white+ expression domain of a mirr enhancer-detector P-element insertion. (E) mirr enhancer-detector expression throughout the eye of a fly in which upd LOF clones have been induced. A slight reduction in eye size and disorder of the ommatidial array in ventral regions is apparent. (F) Section through an eye similar to that shown in E showing the expression domain of the mirr enhancer-detector white+ (dark pigment granules). The approximate area sectioned is indicated by the dotted white box in E. Note that this particular eye had a small region of white− tissue at the ventral edge, which shows normal ventral ommatidial orientation. (G) Schematic representation of F showing the ommatidial orientation with dorsal (green arrow) and ventral (red arrow) fates. Shading indicates the extent of mirr expression (as revealed by regulation of white+). Note that some ommatidia in the mirr-expressing region nevertheless show ventral rotational fate. (H–J) Ventral hop clones do not activate mirr expression. Confocal image of an eye disc containing a ventrally situated hop clone marked by lack of green staining (H,J) also stained for dorsal-specific expression of a mirr enhancer detector in red (I,J).
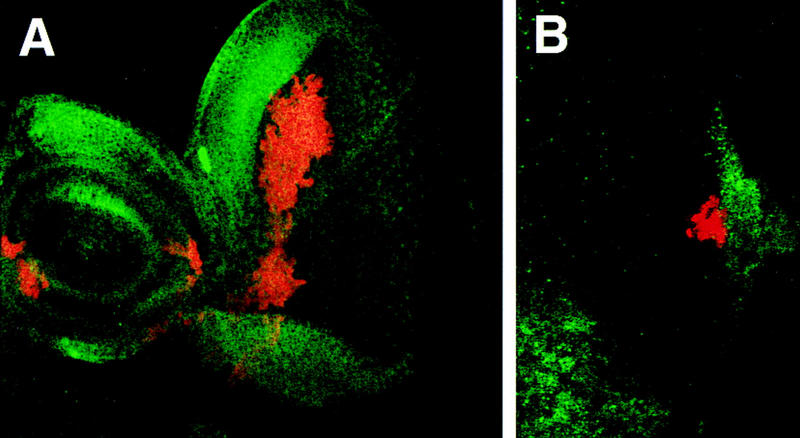
Wg and Upd do not regulate each other’s expression. (A) A disc containing clones of cells ectopically expressing Upd (red), stained for expression of Wg protein (green). Strong Wg expression can be seen at the dorsal margin, adjacent to a large patch of Upd-expressing cells, indicating that Upd does not repress Wg expression. (B) The posterior region of a disc containing clones of cells ectopically expressing Wg (red), also stained for Upd expression (green). Upd expression remains at the optic stalk even when adjacent to Wg-expressing cells.
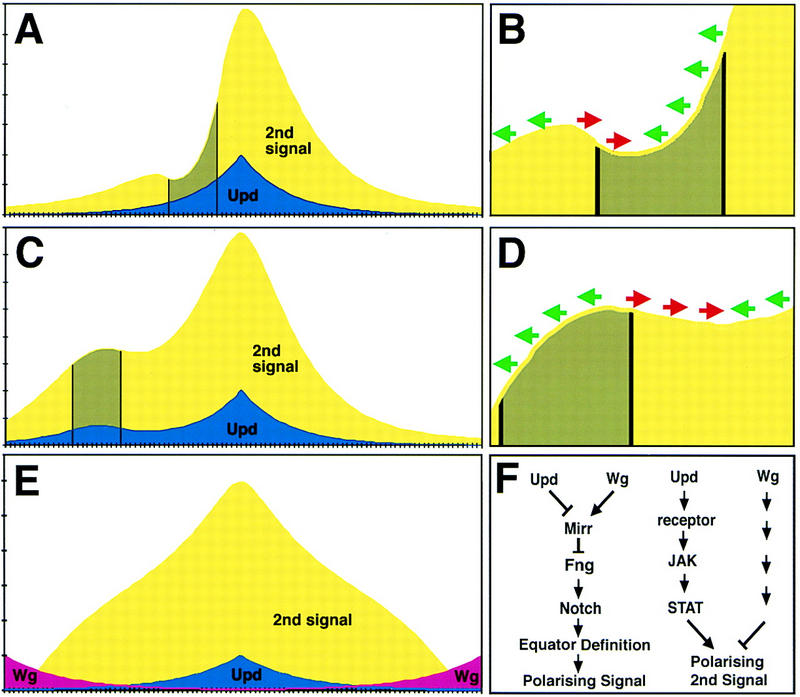
A model for how Upd/JAK/STAT signaling determines ommatidial rotational fate. (A–E) Graphs that represent a single dorsal (left) to ventral (right) line of cells through an eye imaginal disc. The levels of Upd (blue) and second signal (yellow) are shown with the peak of Upd corresponding to the equator or center of the disc. (Green arrows) Dorsal rotational fate; (red arrows) ventral fate (B,D). (A,B) The effect of a JAK/STAT LOF clone (gray) on ommatidial polarity; (C,D) the effect of an Upd misexpressing clone. (E) The predicted concentration of a single second signal if it were both negatively regulated by Wg diffusing from the poles and positively regulated by Upd from the equator; note that the opposing effects of shallow Upd and Wg gradients can combine to produce a steep gradient of second signal. (F) The two proposed functions of Upd/JAK/STAT in ommatidial polarity determination: Upd acts together with Wg to restrict mirr expression and define the position of the equator via activation of Notch, which itself leads to production of an ommatidial polarizing signal; Upd also activates a gradient of JAK/STAT activity that indirectly regulates ommatidial polarity by a mirr-independent mechanism; additionally, Wg secreted from the poles also produces a gradient of an ommatidial polarizing signal.
References
-
- Baker NE, Rubin GM. Ellipse mutations in the Drosophila homologue of the EGF receptor affect pattern formation, cell division, and cell death in eye imaginal discs. Dev Biol. 1992;150:381–396. - PubMed
-
- Binari R, Perrimon N. Stripe-specific regulation of pair-rule genes by hopscotch, a putative Jak family tyrosine kinase in Drosophila. Genes & Dev. 1994;8:300–312. - PubMed
-
- Bonni A, Sun Y, Nadal VM, Bhatt A, Frank DA, Rozovsky I, Stahl N, Yancopoulos GD, Greenberg ME. Regulation of gliagenesis in the central nervous system by the JAK-STAT signaling pathway. Science. 1997;278:477–483. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
-
- Brodsky MH, Steller H. Positional information along the dorsal-ventral axis of the Drosophila eye: Graded expression of the four-jointed gene. Dev Biol. 1996;173:428–446. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases