

Aggregation of bacillus thuringiensis Cry1A toxins upon binding to target insect larval midgut vesicles
- PMID: 10347034
- PMCID: PMC91369
- DOI: 10.1128/AEM.65.6.2503-2507.1999
Aggregation of bacillus thuringiensis Cry1A toxins upon binding to target insect larval midgut vesicles
Abstract
During sporulation, Bacillus thuringiensis produces crystalline inclusions comprised of a mixture of delta-endotoxins. Following ingestion by insect larvae, these inclusion proteins are solubilized, and the protoxins are converted to toxins. These bind specifically to receptors on the surfaces of midgut apical cells and are then incorporated into the membrane to form ion channels. The steps required for toxin insertion into the membrane and possible oligomerization to form a channel have been examined. When bound to vesicles from the midguts of Manduca sexta larvae, the Cry1Ac toxin was largely resistant to digestion with protease K. Only about 60 amino acids were removed from the Cry1Ac amino terminus, which included primarily helix alpha1. Following incubation of the Cry1Ab or Cry1Ac toxins with vesicles, the preparations were solubilized by relatively mild conditions, and the toxin antigens were analyzed by immunoblotting. In both cases, most of the toxin formed a large, antigenic aggregate of ca. 200 kDa. These toxin aggregates did not include the toxin receptor aminopeptidase N, but interactions with other vesicle components were not excluded. No oligomerization occurred when inactive toxins with mutations in amphipathic helices (alpha5) and known to insert into the membrane were tested. Active toxins with other mutations in this helix did form oligomers. There was one exception; a very active helix alpha5 mutant toxin bound very well to membranes, but no oligomers were detected. Toxins with mutations in the loop connecting helices alpha2 and alpha3, which affected the irreversible binding to vesicles, also did not oligomerize. There was a greater extent of oligomerization of the Cry1Ac toxin with vesicles from the Heliothis virescens midgut than with those from the M. sexta midgut, which correlated with observed differences in toxicity. Tight binding of virtually the entire toxin molecule to the membrane and the subsequent oligomerization are both important steps in toxicity.
Figures
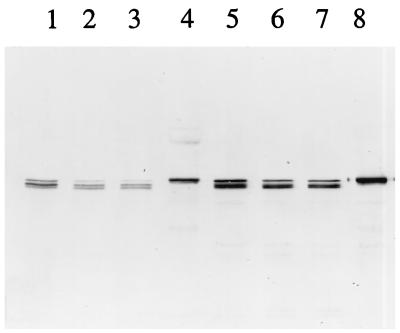
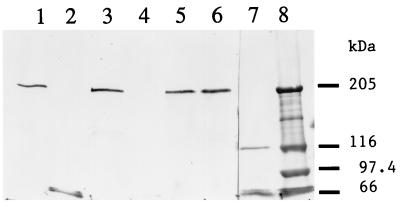
References
-
- Aronson A I. Flexibility in the protoxin composition of Bacillus thuringiensis. FEMS Microbiol Lett. 1994;117:21–28. - PubMed
-
- Carroll J, Convents D, Van Damme J, Boets A, Van Rie J, Ellar D J. Intramolecular proteolytic cleavage of Bacillus thuringiensis Cry3A δ-endotoxin may facilitate its coleopteran toxicity. J Invertebr Pathol. 1997;70:41–49. - PubMed
-
- Chen X, Curtiss A, Alcantra E, Dean D. Mutations in domain 1 of the Bacillus thuringiensis δ-endotoxin CryIAb reduce the irreversible binding of toxin to Manduca sexta brush border membrane vesicles. J Biol Chem. 1995;270:6412–6419. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
