

Cell cycle expression and transcriptional regulation of DNA topoisomerase IV genes in caulobacter
- PMID: 10348842
- PMCID: PMC93797
- DOI: 10.1128/JB.181.11.3321-3329.1999
Cell cycle expression and transcriptional regulation of DNA topoisomerase IV genes in caulobacter
Abstract
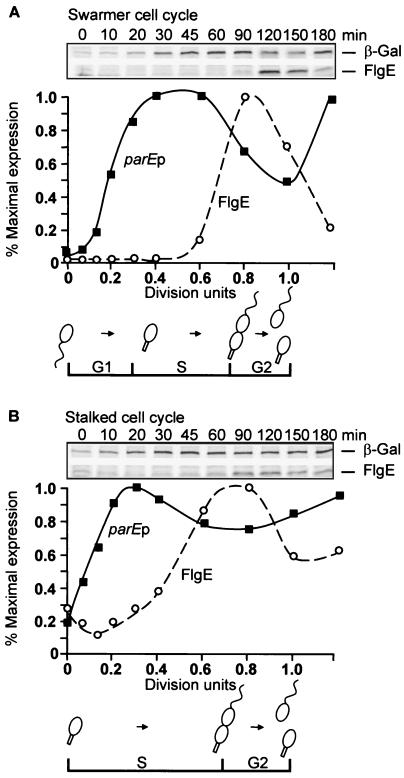
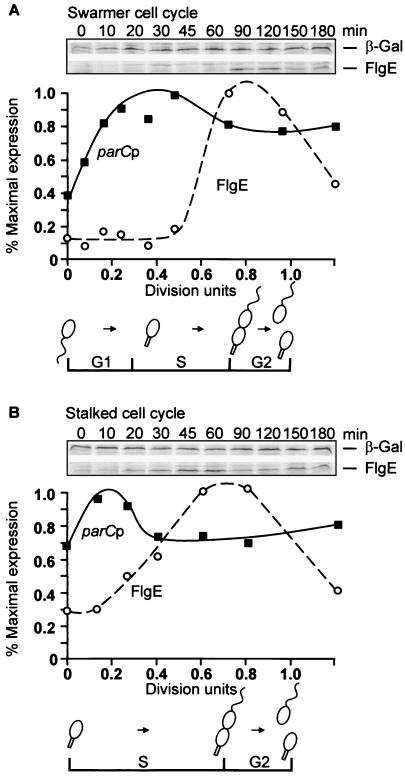
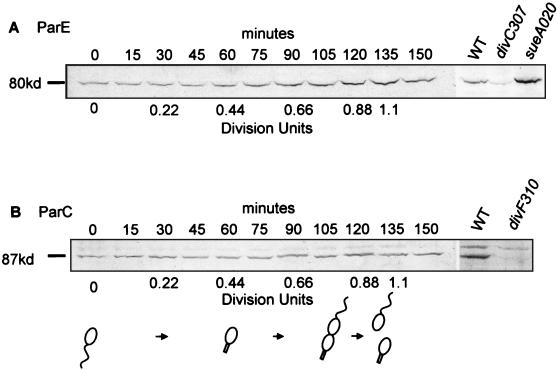
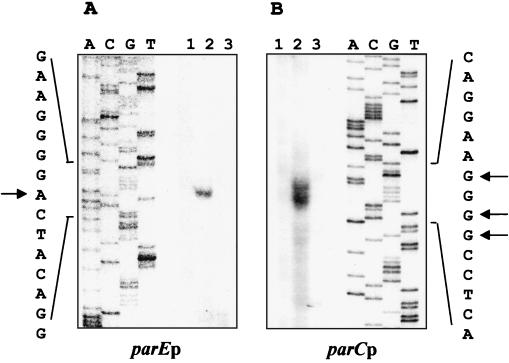
DNA replication and differentiation are closely coupled during the Caulobacter crescentus cell cycle. We have previously shown that DNA topoisomerase IV (topo IV), which is encoded by the parE and parC genes, is required for chromosomal partitioning, cell division, and differentiation in this bacterium (D. Ward and A. Newton, Mol. Microbiol. 26:897-910, 1997). We have examined the cell cycle regulation of parE and parC and report here that transcription of these topo IV genes is induced during the swarmer-to-stalked-cell transition when cells prepare for initiation of DNA synthesis. The regulation of parE and parC expression is not strictly coordinated, however. The rate of parE transcription increases ca. 20-fold during the G1-to-S-phase transition and in this respect, its pattern of regulation is similar to those of several other genes required for chromosome duplication. Transcription from the parC promoter, by contrast, is induced only two- to threefold during this cell cycle period. Steady-state ParE levels are also regulated, increasing ca. twofold from low levels in swarmer cells to a maximum immediately prior to cell division, while differences in ParC levels during the cell cycle could not be detected. These results suggest that topo IV activity may be regulated primarily through parE expression. The presumptive promoters of the topo IV genes display striking similarities to, as well as differences from, the consensus promoter recognized by the major Caulobacter sigma factor sigma73. We also present evidence that a conserved 8-mer sequence motif located in the spacers between the -10 and -35 elements of the parE and parC promoters is required for maximum levels of parE transcription, which raises the possibility that it may function as a positive regulatory element. The pattern of parE transcription and the parE and parC promoter architecture suggest that the topo IV genes belong to a specialized subset of cell cycle-regulated genes required for chromosome replication.
Figures


Similar articles
-
Requirement of topoisomerase IV parC and parE genes for cell cycle progression and developmental regulation in Caulobacter crescentus.Mol Microbiol. 1997 Dec;26(5):897-910. doi: 10.1046/j.1365-2958.1997.6242005.x. Mol Microbiol. 1997. PMID: 9426128
-
Transcription of genes encoding DNA replication proteins is coincident with cell cycle control of DNA replication in Caulobacter crescentus.J Bacteriol. 1997 Apr;179(7):2319-30. doi: 10.1128/jb.179.7.2319-2330.1997. J Bacteriol. 1997. PMID: 9079919 Free PMC article.
-
Conserved promoter motif is required for cell cycle timing of dnaX transcription in Caulobacter.J Bacteriol. 2001 Aug;183(16):4860-5. doi: 10.1128/JB.183.16.4860-4865.2001. J Bacteriol. 2001. PMID: 11466289 Free PMC article.
-
Regulation of cellular differentiation in Caulobacter crescentus.Microbiol Rev. 1995 Mar;59(1):31-47. doi: 10.1128/mr.59.1.31-47.1995. Microbiol Rev. 1995. PMID: 7708011 Free PMC article. Review.
-
Cis- and trans-acting elements required for regulation of flagellar gene transcription in the bacterium Caulobacter crescentus.Cell Mol Biol Res. 1993;39(4):361-9. Cell Mol Biol Res. 1993. PMID: 8312972 Review.
Cited by
-
The regulation of DNA supercoiling across evolution.Protein Sci. 2021 Oct;30(10):2042-2056. doi: 10.1002/pro.4171. Epub 2021 Aug 23. Protein Sci. 2021. PMID: 34398513 Free PMC article. Review.
-
The topoisomerase IV ParC subunit colocalizes with the Caulobacter replisome and is required for polar localization of replication origins.Proc Natl Acad Sci U S A. 2004 Jun 22;101(25):9251-6. doi: 10.1073/pnas.0402567101. Epub 2004 Jun 3. Proc Natl Acad Sci U S A. 2004. PMID: 15178756 Free PMC article.
References
-
- Ausubel F M, Brent R, Kingston R, Moore D, editors. Short protocols in molecular biology. 3rd ed. New York, N.Y: John Wiley and Sons, Inc.; 1995.
-
- Berk J A, Sharp P A. Sizing and mapping of early adenovirus mRNAs by gel electrophoresis of S1 endonuclease-digested hybrids. Cell. 1977;12:721–732. - PubMed
-
- Brun Y V, Marczynski G, Shapiro L. The expression of asymmetry during Caulobacter cell differentiation. Annu Rev Biochem. 1994;63:419–450. - PubMed
-
- Ching Y, Dinh T, Beran R K. Multiple promoters for transcription of the Escherichia coli DNA topoisomerase I gene and their regulation by DNA supercoiling. J Mol Biol. 1988;202:735–742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
