

Early expression of the calmodulin gene, which precedes appressorium formation in Magnaporthe grisea, is inhibited by self-inhibitors and requires surface attachment
- PMID: 10348871
- PMCID: PMC93826
- DOI: 10.1128/JB.181.11.3571-3577.1999
Early expression of the calmodulin gene, which precedes appressorium formation in Magnaporthe grisea, is inhibited by self-inhibitors and requires surface attachment
Abstract
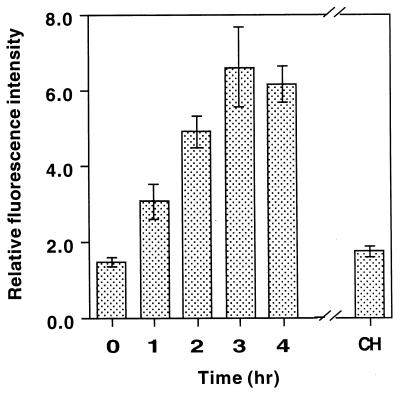
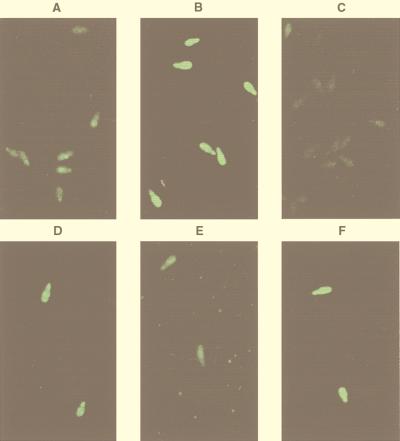
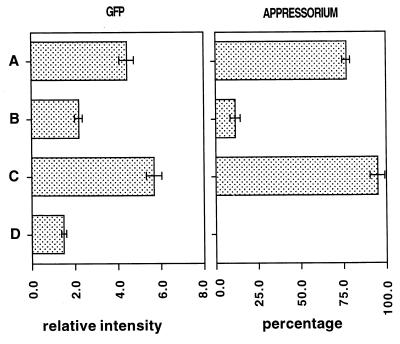
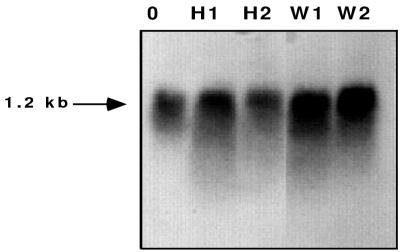
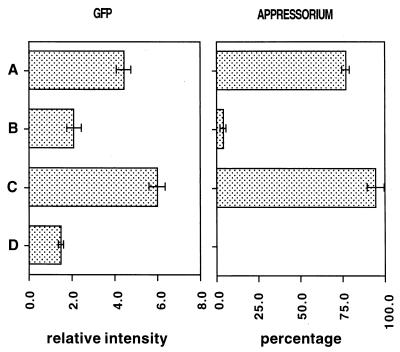
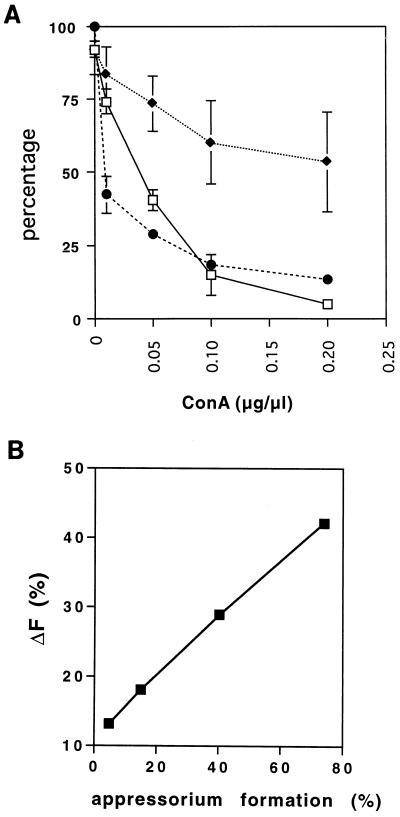
Fungal conidia contain chemicals that inhibit germination and appressorium formation until they are well dispersed in a favorable environment. Recently, such self-inhibitors were found to be present on the conidia of Magnaporthe grisea, and plant surface waxes were found to relieve this self-inhibition. To determine whether the self-inhibitors suppress the expression of early genes involved in the germination and differentiation of conidia, the calmodulin gene was chosen as a representative early gene, because it was found to be expressed early in Colletotrichum gloeosporioides and Colletotrichum trifolii differentiation. After calmodulin cDNA and genomic DNA from M. grisea were cloned, the promoter of the calmodulin gene was fused to a reporter gene, that for green fluorescent protein (GFP), and transformed into the M. grisea genome. Confocal microscopic examination and quantitation of expression of GFP green fluorescence showed (i) that the expression of the calmodulin gene decreased significantly when self-inhibition of M. grisea appressorium formation occurred because of high conidial density or addition of exogenous self-inhibitors and (ii) that the expression level of this gene was restored when self-inhibition was relieved by the addition of plant surface waxes. The increase in fluorescence correlated with the percentage of conidia that formed appressoria. The induction of calmodulin was also confirmed by RNA blotting. Concanavalin A inhibited surface attachment of conidia, GFP expression, and appressorium formation without affecting germination. The high correlation between GFP expression and appressorium formation strongly suggests that calmodulin gene expression and appressorium formation require surface attachment.
Figures
Similar articles
-
Induction of Ca2+-calmodulin signaling by hard-surface contact primes Colletotrichum gloeosporioides conidia to germinate and form appressoria.J Bacteriol. 1998 Oct;180(19):5144-50. doi: 10.1128/JB.180.19.5144-5150.1998. J Bacteriol. 1998. PMID: 9748448 Free PMC article.
-
Regulation of the MPG1 hydrophobin gene in the rice blast fungus Magnaporthe grisea.Mol Plant Microbe Interact. 2002 Dec;15(12):1253-67. doi: 10.1094/MPMI.2002.15.12.1253. Mol Plant Microbe Interact. 2002. PMID: 12481998
-
Bypassing both surface attachment and surface recognition requirements for appressorium formation by overactive ras signaling in Magnaporthe oryzae.Mol Plant Microbe Interact. 2014 Sep;27(9):996-1004. doi: 10.1094/MPMI-02-14-0052-R. Mol Plant Microbe Interact. 2014. PMID: 24835254
-
Serial Analysis of Gene Expression (SAGE) of Magnaporthe grisea: genes involved in appressorium formation.Mol Genet Genomics. 2003 Nov;270(2):181-9. doi: 10.1007/s00438-003-0911-6. Epub 2003 Aug 29. Mol Genet Genomics. 2003. PMID: 12955499
-
Infection-related development in the rice blast fungus Magnaporthe grisea.Curr Opin Microbiol. 1998 Dec;1(6):693-7. doi: 10.1016/s1369-5274(98)80117-3. Curr Opin Microbiol. 1998. PMID: 10066544 Review.
Cited by
-
Isolation of mycoparasitic-related transcripts by SSH during interaction of the mycoparasite Stachybotrys elegans with its host Rhizoctonia solani.Curr Genet. 2008 Feb;53(2):67-80. doi: 10.1007/s00294-007-0166-6. Epub 2007 Dec 5. Curr Genet. 2008. PMID: 18058103
-
Nonpathogenic strains of Colletotrichum lindemuthianum trigger progressive bean defense responses during appressorium-mediated penetration.Appl Environ Microbiol. 2005 Aug;71(8):4761-70. doi: 10.1128/AEM.71.8.4761-4770.2005. Appl Environ Microbiol. 2005. PMID: 16085873 Free PMC article.
-
Hyphal Fusions Enable Efficient Nutrient Distribution in Colletotrichum graminicola Conidiation and Symptom Development on Maize.Microorganisms. 2022 Jun 1;10(6):1146. doi: 10.3390/microorganisms10061146. Microorganisms. 2022. PMID: 35744664 Free PMC article.
-
Friends or foes? Emerging insights from fungal interactions with plants.FEMS Microbiol Rev. 2016 Mar;40(2):182-207. doi: 10.1093/femsre/fuv045. Epub 2015 Nov 21. FEMS Microbiol Rev. 2016. PMID: 26591004 Free PMC article. Review.
-
Green Fluorescent Protein Expression in Pseudogymnoascus destructans to Study Its Abiotic and Biotic Lifestyles.Mycopathologia. 2018 Oct;183(5):805-814. doi: 10.1007/s11046-018-0285-2. Epub 2018 Jul 9. Mycopathologia. 2018. PMID: 29987576
References
-
- Braam J, Davis R W. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell. 1990;60:357–364. - PubMed
-
- Cormack B P, Valdivia R H, Falkow S. FACS-optimized mutants of the green fluorescent protein (GFP) Gene. 1996;173:33–38. - PubMed
-
- Dean R A. Signal pathways and appressorium morphogenesis. Annu Rev Phytopathol. 1997;35:211–234. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
