

doi: 10.1128/JB.181.11.3587-3590.1999.
Importance of a 5' stem-loop for longevity of papA mRNA in Escherichia coli
Affiliations
- PMID: 10348874
- PMCID: PMC93829
- DOI: 10.1128/JB.181.11.3587-3590.1999
Item in Clipboard
Importance of a 5' stem-loop for longevity of papA mRNA in Escherichia coli
J Bacteriol.
1999 Jun.
Abstract
High-level expression of the major pilus subunit (PapA) of uropathogenic strains of Escherichia coli results in part from the unusually long lifetime of the mRNA that encodes this protein. Here we report that the longevity of papA mRNA derives in large measure from the protection afforded by its 5' untranslated region. This papA RNA segment can prolong the lifetime of an otherwise short-lived mRNA to which it is fused. In vivo alkylation studies indicate that, in its natural milieu, the papA message begins with a stem-loop structure. This stem-loop is important for the stabilizing effect of the papA 5' untranslated region, as evidenced by the significant acceleration in papA mRNA decay that results from its removal.
Figures
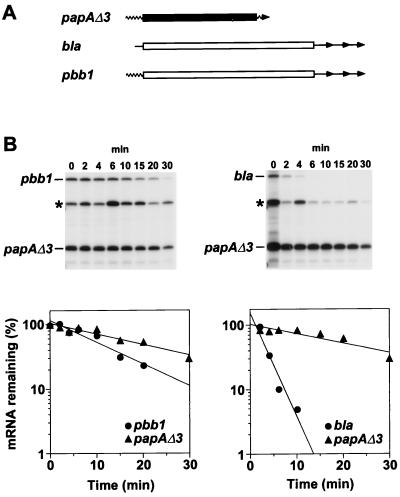
Decay of a hybrid papA-bla mRNA. (A) The papA-bla hybrid transcript pbb1 is represented diagrammatically along with its mRNA progenitors papAΔ3 and bla. These mRNAs were expressed from plasmid pPBB1E or pBLAE, each a derivative of plasmid pBLA200 (8), by transcription from a bla promoter. Wavy lines, papA UTRs; straight lines, bla UTRs; solid rectangle, papA coding region: open rectangles, bla coding region; arrowheads, mRNA 3′ ends. There are three alternative sites of transcription termination downstream of the bla gene. (B) Cultures of a recA mutant derivative of E. coli MG1693 (2) harboring either pPBB1E (left) or pBLAE (right) were grown exponentially for several generations at 37°C in Luria-Bertani medium supplemented with glucose (0.4%) and Casamino Acids (0.5%). At time intervals after transcription inhibition with rifampin (200 μg/ml), total cellular RNA was isolated. Equal amounts of each RNA sample (2 μg) were then analyzed by S1 protection with a mixture of two 5′-end-labeled probes complementary to a 5′-terminal segment of papAΔ3 mRNA or bla mRNA. The radioactivity in bands that correspond to papAΔ3, pbb1, and bla mRNA was quantitated with a Molecular Dynamics PhosphorImager. Also marked is a band corresponding to the reannealed papA probe DNA (∗). Beneath each autoradiogram is a semilogarithmic plot of mRNA concentration versus time. Half-lives were calculated from the slope of each plot, and half-life errors were estimated from the standard deviation of the slopes. The measured half-lives were 8.9 ± 1.1 min for pbb1 mRNA and 18 ± 2 min for papAΔ3 mRNA (left) and 1.9 ± 0.4 min for bla mRNA and 20 ± 4 min for papAΔ3 mRNA (right). For procedural details concerning bacterial cell growth, RNA isolation, and S1 analysis, see reference .
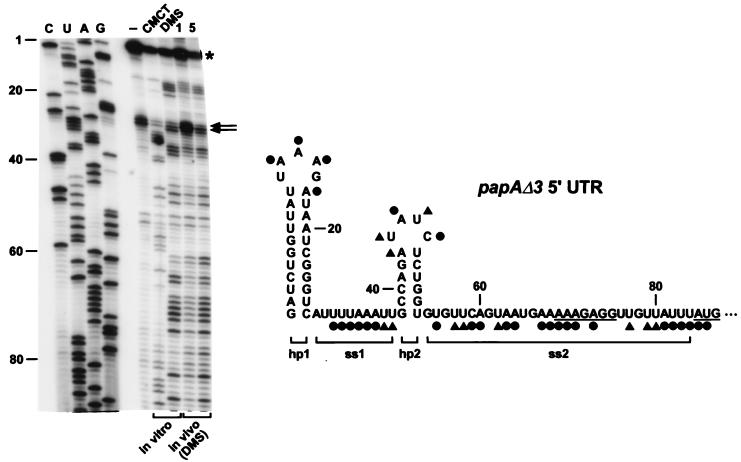
Alkylation of the 5′ UTR of papAΔ3 mRNA. (Left) Primer extension analysis of alkylated papAΔ3 mRNA. Total cellular RNA was isolated from an exponential-phase culture of E. coli C600S (17) containing pPAPAΔ3 after treatment of aliquots of the culture with DMS (in vivo, 1 or 5 μl per ml of culture). In addition, samples of RNA extracted from an untreated culture were alkylated in vitro with DMS or CMCT. Sites of alkylation were mapped by primer extension with avian myeloblastosis virus reverse transcriptase and a 5′-end-labeled DNA primer (5′-AAGACACCACTGCCATAGCT-3′) complementary to the coding region of papA mRNA. The resulting extension products were then analyzed by gel electrophoresis beside sequencing ladders that were generated by extension of the same 5′-end-labeled primer on a papAΔ3 DNA template. Unalkylated RNA (lane −) served as a negative control to identify primer extension products unrelated to alkylation. Blockage of primer extension by an alkylated RNA base results in the production of a complementary DNA fragment one nucleotide shorter than that arising from incorporation of a dideoxynucleotide opposite the same base. In the experiment shown, CMCT did not react detectably with guanosine nucleotides, precluding a direct assessment of base pairing by those residues. The sequencing lanes (C, U, A, G) are labeled to indicate the sequence of the RNA, not the complementary DNA. Calibration is in nucleotides from the papAΔ3 5′ end. An asterisk marks the site of transcription initiation. The degree of chemical modification can be difficult to assess at sites where it is no greater than the basal level of termination by reverse transcriptase on an unalkylated RNA template. With the avian myeloblastosis virus enzyme, such sites often correspond to the 3′ boundary of secondary structure elements of significant thermodynamic stability (6), and in this case, the two major sites of premature termination on unalkylated RNA (marked by arrows) map to the foot of the 5′-terminal papAΔ3 stem-loop. For procedural details, see reference . (Right) Summary of the alkylation data for the papAΔ3 5′ UTR. ●, heavy alkylation; ▴, moderate alkylation. Brackets delineate the boundaries of the four structural domains (hp1, ss1, hp2, and ss2) within the papAΔ3 5′ UTR. The Shine-Dalgarno element and initiation codon are underlined.
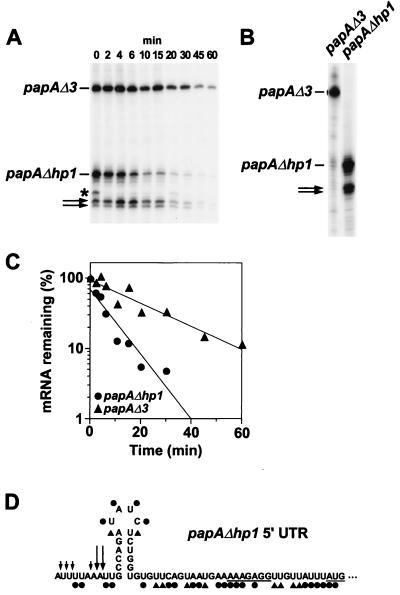
Accelerated decay of a truncated papA transcript lacking the 5′ stem-loop. (A) A culture of E. coli JC10287 (7) harboring pPAPAΔhp1E was grown exponentially for several generations in supplemented Luria-Bertani medium at 37°C. At time intervals after transcription inhibition with rifampin (200 μg/ml), total cellular RNA was isolated. Equal amounts of each RNA sample (2 μg) were then analyzed by primer extension with avian myeloblastosis virus reverse transcriptase and a 5′-end-labeled DNA primer (5′-AAGACACCACTGCCATAGCT-3′) complementary to the coding region of papA mRNA. Bands that correspond to papAΔ3 and papAΔhp1 mRNA are indicated, as are two bands corresponding to a pair of apparent papA mRNA cleavage products (arrows). The origin of an additional band (∗) seen only at 0 min is not known. (B) That the two cleavage products are derived principally from papAΔhp1 mRNA and not significantly from papA mRNA is demonstrated by an additional primer extension experiment performed with RNA samples isolated from a pair of isogenic E. coli strains that expressed either papAΔ3 mRNA or papAΔhp1 mRNA. (C) Semilogarithmic plot of mRNA concentration versus time after rifampin addition. The measured half-lives were 6.6 ± 0.9 min for papAΔhp1 mRNA and 18 ± 2 min for papAΔ3 mRNA. (D) Summary of the DMS and CMCT alkylation data obtained in vivo and in vitro for the 5′ UTR of papAΔhp1 mRNA by the procedure described in Fig. 2. ●, heavy alkylation; ▴, moderate alkylation. Sites of premature termination by reverse transcriptase on an unalkylated RNA template are indicated by arrows: large arrows, major termination sites thought to represent the 5′ ends of RNase E cleavage products generated in vivo; small arrows, minor termination sites that preclude a direct assessment of base pairing by the preceding residue. The Shine-Dalgarno element and initiation codon are underlined.
Similar articles
-
An evolutionarily conserved RNA stem-loop functions as a sensor that directs feedback regulation of RNase E gene expression.Genes Dev. 2000 May 15;14(10):1249-60. Genes Dev. 2000. PMID: 10817759 Free PMC article.
-
A conserved RNA structure (thi box) is involved in regulation of thiamin biosynthetic gene expression in bacteria.Proc Natl Acad Sci U S A. 2001 Aug 14;98(17):9736-41. doi: 10.1073/pnas.161168098. Epub 2001 Jul 24. Proc Natl Acad Sci U S A. 2001. PMID: 11470904 Free PMC article.
-
A 5' stem-loop and ribosome binding but not translation are important for the stability of Bacillus subtilis aprE leader mRNA.Microbiology (Reading). 2002 Jun;148(Pt 6):1795-1803. doi: 10.1099/00221287-148-6-1795. Microbiology (Reading). 2002. PMID: 12055299
-
Translation initiation and the fate of bacterial mRNAs.FEMS Microbiol Rev. 2006 Nov;30(6):967-79. doi: 10.1111/j.1574-6976.2006.00043.x. Epub 2006 Sep 21. FEMS Microbiol Rev. 2006. PMID: 16989654 Review.
-
Measuring the immeasurable.Mol Cell. 2002 Sep;10(3):437-9. doi: 10.1016/s1097-2765(02)00661-5. Mol Cell. 2002. PMID: 12408812 Review.
Cited by
-
Adjacent single-stranded regions mediate processing of tRNA precursors by RNase E direct entry.Nucleic Acids Res. 2014 Apr;42(7):4577-89. doi: 10.1093/nar/gkt1403. Epub 2014 Jan 21. Nucleic Acids Res. 2014. PMID: 24452799 Free PMC article.
-
A VapBC toxin-antitoxin module is a posttranscriptional regulator of metabolic flux in mycobacteria.J Bacteriol. 2012 May;194(9):2189-204. doi: 10.1128/JB.06790-11. Epub 2012 Feb 24. J Bacteriol. 2012. PMID: 22366418 Free PMC article.
-
The Bacillus subtilis late competence operon comE is transcriptionally regulated by yutB and under post-transcription initiation control by comN (yrzD).J Bacteriol. 2009 Feb;191(3):949-58. doi: 10.1128/JB.01429-08. Epub 2008 Nov 21. J Bacteriol. 2009. PMID: 19028902 Free PMC article.
-
Initiation of RNA decay in Escherichia coli by 5' pyrophosphate removal.Mol Cell. 2007 Jul 6;27(1):79-90. doi: 10.1016/j.molcel.2007.05.038. Mol Cell. 2007. PMID: 17612492 Free PMC article.
-
Regulation of clpQ⁺Y⁺ (hslV⁺U⁺) gene expression in Escherichia coli.Open Microbiol J. 2009;3:29-39. doi: 10.2174/1874285800903010029. Epub 2009 Mar 17. Open Microbiol J. 2009. PMID: 19440251 Free PMC article.
References
-
- Båga M, Göransson M, Normark S, Uhlin B E. Processed mRNA with differential stability in the regulation of E. coli pilin gene expression. Cell. 1988;52:197–206. - PubMed
-
- Belasco J G, Nilsson G, von Gabain A, Cohen S N. The stability of E. coli gene transcripts is dependent on determinants localized to specific mRNA segments. Cell. 1986;46:245–251. - PubMed
-
- Bouvet P, Belasco J G. Control of RNase E-mediated RNA degradation by 5′-terminal base pairing in E. coli. Nature. 1992;360:488–491. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
