

Separate effects of a classical conditioning procedure on respiratory pumping, swimming, and inking in Aplysia fasciata
- PMID: 10355521
- PMCID: PMC311277
Separate effects of a classical conditioning procedure on respiratory pumping, swimming, and inking in Aplysia fasciata
Abstract
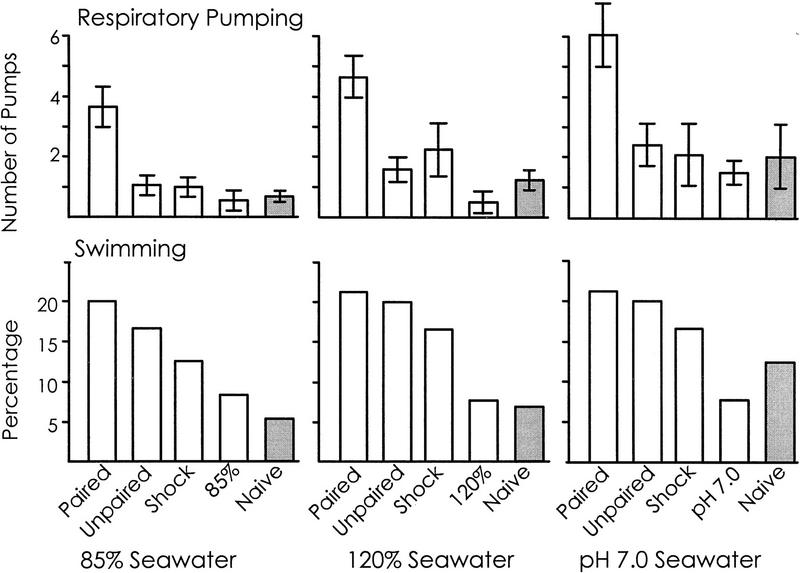
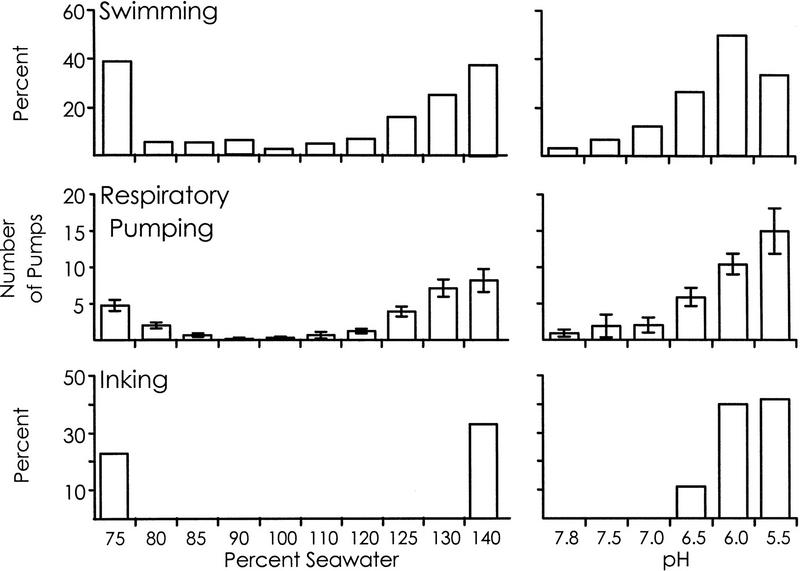
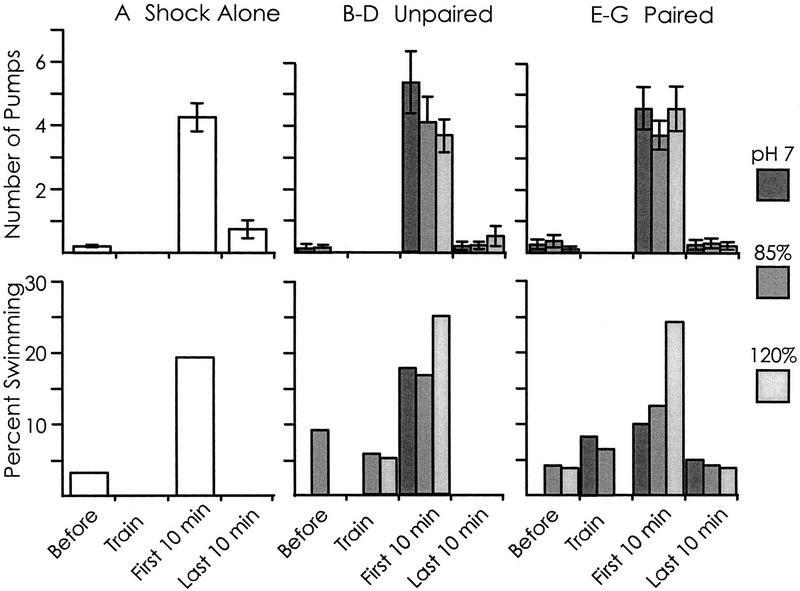
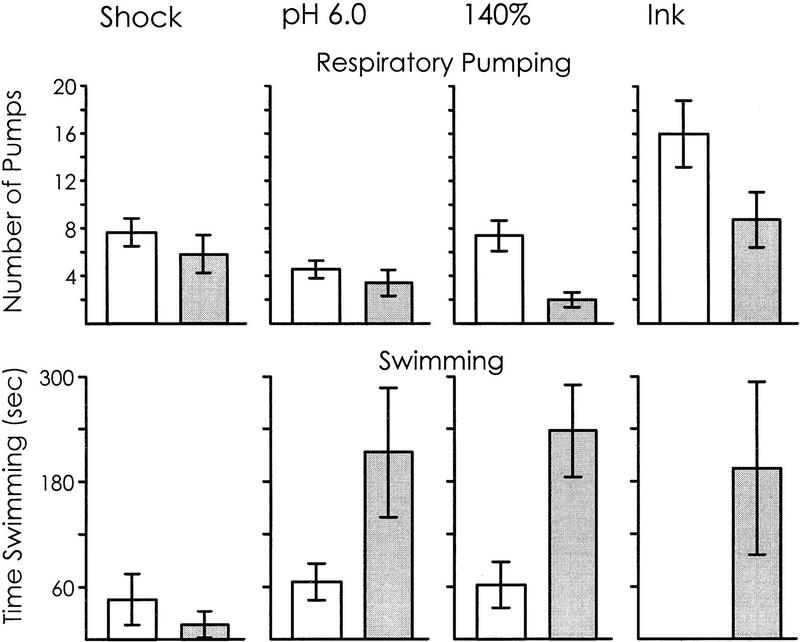
We examined whether swimming and inking, two defensive responses in Aplysia fasciata, are facilitated by a classical conditioning procedure that has been shown to facilitate a third defensive response, respiratory pumping. Training consisted of pairing a head shock (UCS) with a modified seawater (85%, 120%, or pH 7.0 seawater--CSs). Animals were tested by re-exposing them to the same altered seawater 1 hr after the training. For all three altered seawaters, only respiratory pumping is specifically increased by conditioning. Swimming is sensitized by shock, and inking is unaffected by training, indicating that the conditioning procedure is likely to affect a neural site that differentially controls respiratory pumping. Additional observations also indicate that the three defensive responses are differentially regulated. First, different noxious stimuli preferentially elicit different defensive responses. Second, the three defensive responses are differentially affected by shock. Inking is elicited only immediately following shock, whereas swimming and respiratory pumping are facilitated for a period of time following the shock. Third, swimming and respiratory pumping are differentially affected by noxious stimuli that are delivered in open versus closed environments. These data confirm that neural pathways exist that allow Aplysia to modulate separately each of the three defensive behaviors that were examined.
Figures

Similar articles
-
Learned changes in the rate of respiratory pumping in Aplysia fasciata in response to increases and decreases in seawater concentration.Behav Neurosci. 1994 Feb;108(1):161-70. doi: 10.1037//0735-7044.108.1.161. Behav Neurosci. 1994. PMID: 8192842
-
Separate neural pathways respond to different noxious stimuli affecting respiratory pump frequency in Aplysia fasciata.Brain Res. 1993 Jul 9;616(1-2):218-29. doi: 10.1016/0006-8993(93)90212-6. Brain Res. 1993. PMID: 8358614
-
Learned changes of respiratory pump rate in response to lowered pH in Aplysia.Behav Neural Biol. 1990 Nov;54(3):218-33. doi: 10.1016/0163-1047(90)90606-7. Behav Neural Biol. 1990. PMID: 2078160
-
Roles of second messenger pathways in neuronal plasticity and in learning and memory. Insights gained from Aplysia.Adv Second Messenger Phosphoprotein Res. 1993;27:47-108. Adv Second Messenger Phosphoprotein Res. 1993. PMID: 8418826 Review. No abstract available.
-
Feeding behavior of Aplysia: a model system for comparing cellular mechanisms of classical and operant conditioning.Learn Mem. 2006 Nov-Dec;13(6):669-80. doi: 10.1101/lm.339206. Learn Mem. 2006. PMID: 17142299 Review.
Cited by
-
Evolutionary Developmental Biology and Human Language Evolution: Constraints on Adaptation.Evol Biol. 2012 Dec;39(4):613-637. doi: 10.1007/s11692-012-9162-y. Epub 2012 Mar 7. Evol Biol. 2012. PMID: 23226905 Free PMC article.
-
Breathing: rhythmicity, plasticity, chemosensitivity.Annu Rev Neurosci. 2003;26:239-66. doi: 10.1146/annurev.neuro.26.041002.131103. Epub 2003 Feb 13. Annu Rev Neurosci. 2003. PMID: 12598679 Free PMC article. Review.
-
Rapid and persistent suppression of feeding behavior induced by sensitization training in Aplysia.Learn Mem. 2012 Mar 14;19(4):159-63. doi: 10.1101/lm.024638.111. Learn Mem. 2012. PMID: 22419814 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources