

Structural features of the glutamate transporter family
- PMID: 10357852
- PMCID: PMC98967
- DOI: 10.1128/MMBR.63.2.293-307.1999
Structural features of the glutamate transporter family
Abstract
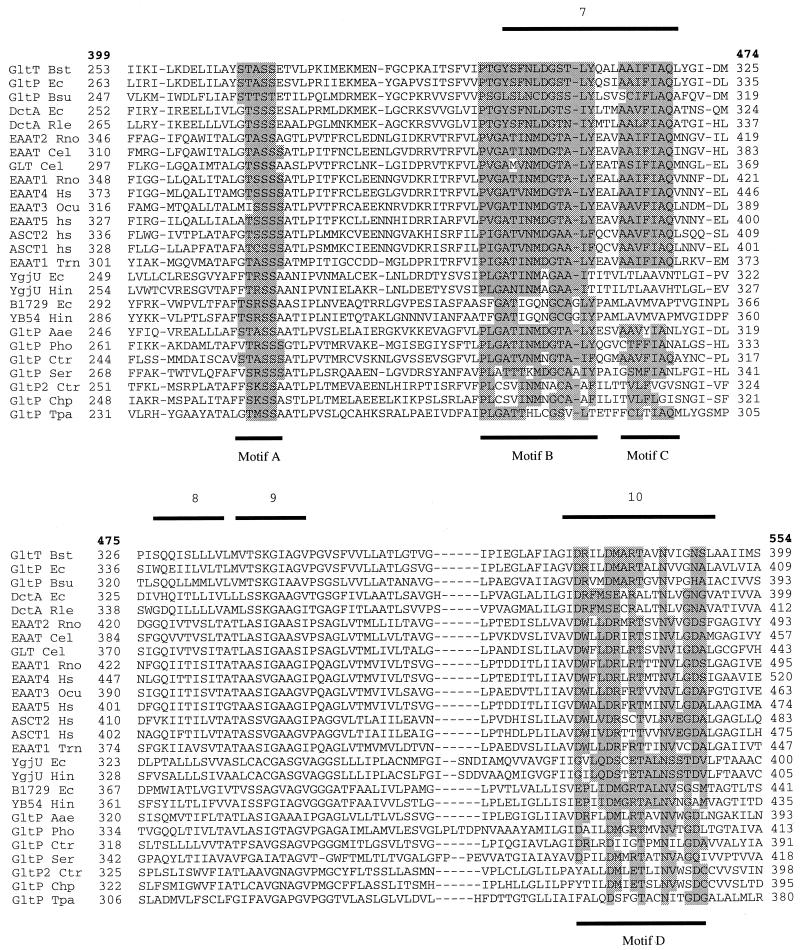
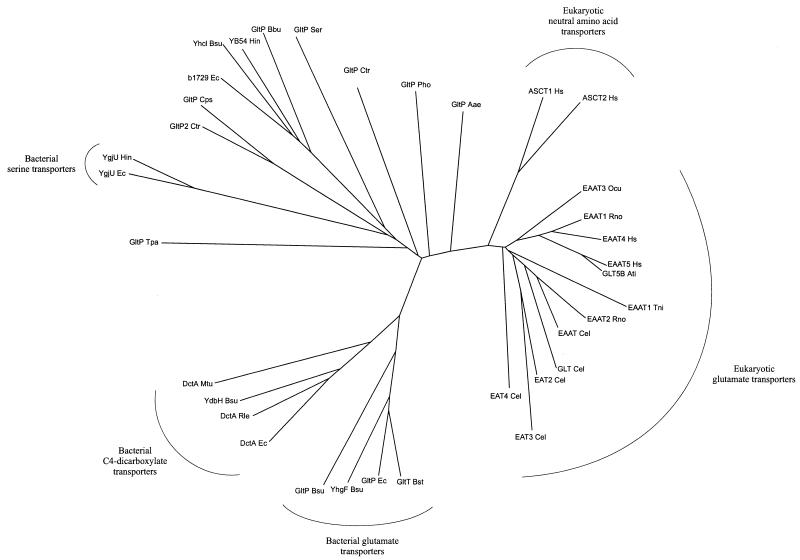
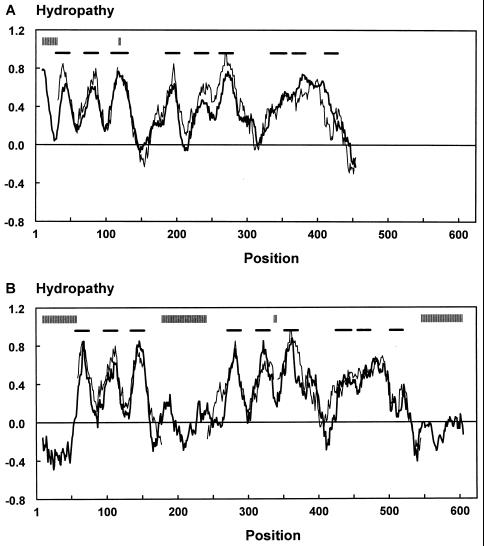
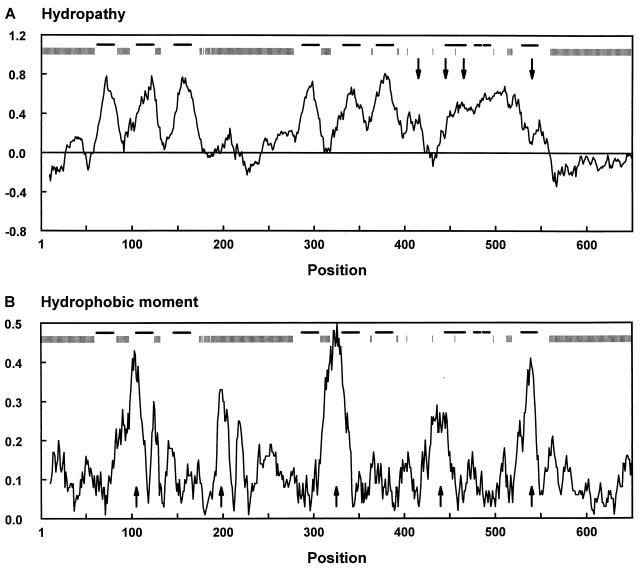
Neuronal and glial glutamate transporters remove the excitatory neurotransmitter glutamate from the synaptic cleft and thus prevent neurotoxicity. The proteins belong to a large and widespread family of secondary transporters, including bacterial glutamate, serine, and C4-dicarboxylate transporters; mammalian neutral-amino-acid transporters; and an increasing number of bacterial, archaeal, and eukaryotic proteins that have not yet been functionally characterized. Sixty members of the glutamate transporter family were found in the databases on the basis of sequence homology. The amino acid sequences of the carriers have diverged enormously. Homology between the members of the family is most apparent in a stretch of approximately 150 residues in the C-terminal part of the proteins. This region contains four reasonably well-conserved sequence motifs, all of which have been suggested to be part of the translocation pore or substrate binding site. Phylogenetic analysis of the C-terminal stretch revealed the presence of five subfamilies with characterized members: (i) the eukaryotic glutamate transporters, (ii) the bacterial glutamate transporters, (iii) the eukaryotic neutral-amino-acid transporters, (iv) the bacterial C4-dicarboxylate transporters, and (v) the bacterial serine transporters. A number of other subfamilies that do not contain characterized members have been defined. In contrast to their amino acid sequences, the hydropathy profiles of the members of the family are extremely well conserved. Analysis of the hydropathy profiles has suggested that the glutamate transporters have a global structure that is unique among secondary transporters. Experimentally, the unique structure of the transporters was recently confirmed by membrane topology studies. Although there is still controversy about part of the topology, the most likely model predicts the presence of eight membrane-spanning alpha-helices and a loop-pore structure which is unique among secondary transporters but may resemble loop-pores found in ion channels. A second distinctive structural feature is the presence of a highly amphipathic membrane-spanning helix that provides a hydrophilic path through the membrane. Recent data from analysis of site-directed mutants and studies on the mechanism and pharmacology of the transporters are discussed in relation to the structural model.
Figures
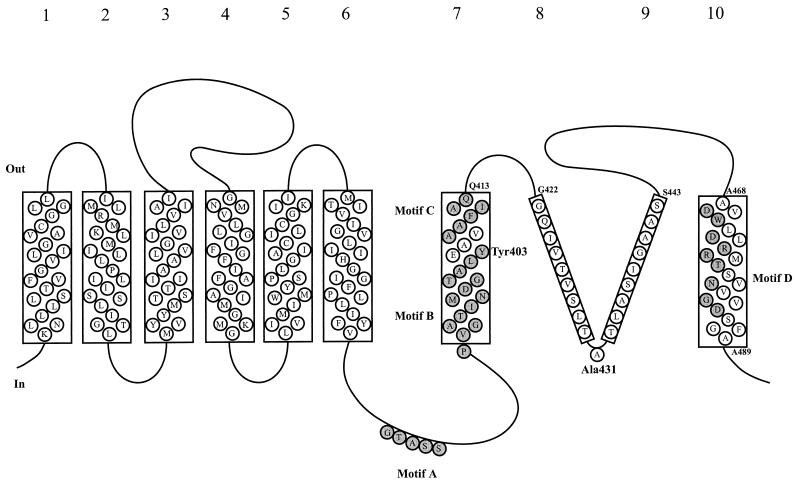
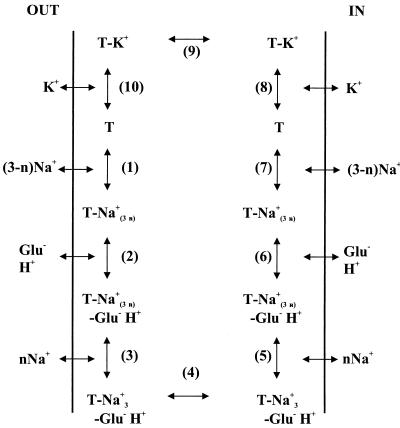
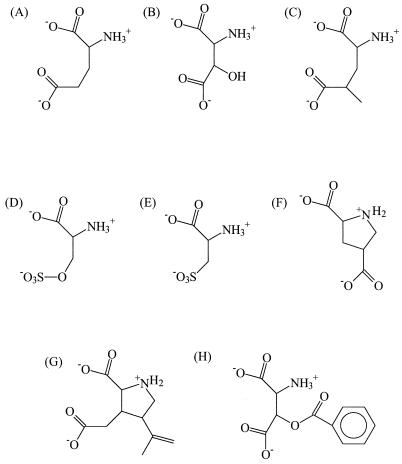
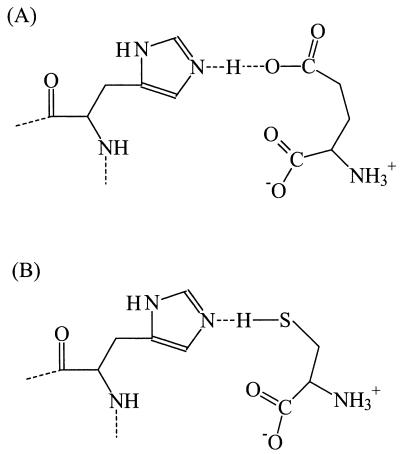
References
-
- Arriza J L, Kavanaugh M P, Fairman W A, Wu Y N, Murdoch G H, North R A, Amara S G. Cloning and expression of a human neutral amino acid transporter with structural similarity to the glutamate transporter gene family. J Biol Chem. 1993;268:15329–15332. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
