

Pharmacological evidence for a KATP channel in renin-secreting cells from rat kidney
- PMID: 10358118
- PMCID: PMC2269373
- DOI: 10.1111/j.1469-7793.1999.0781s.x
Pharmacological evidence for a KATP channel in renin-secreting cells from rat kidney
Abstract
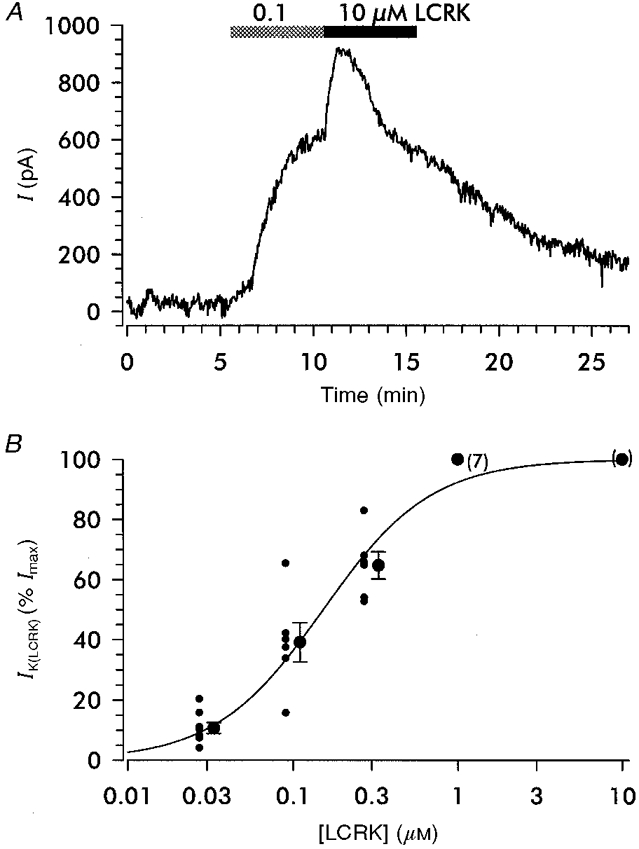
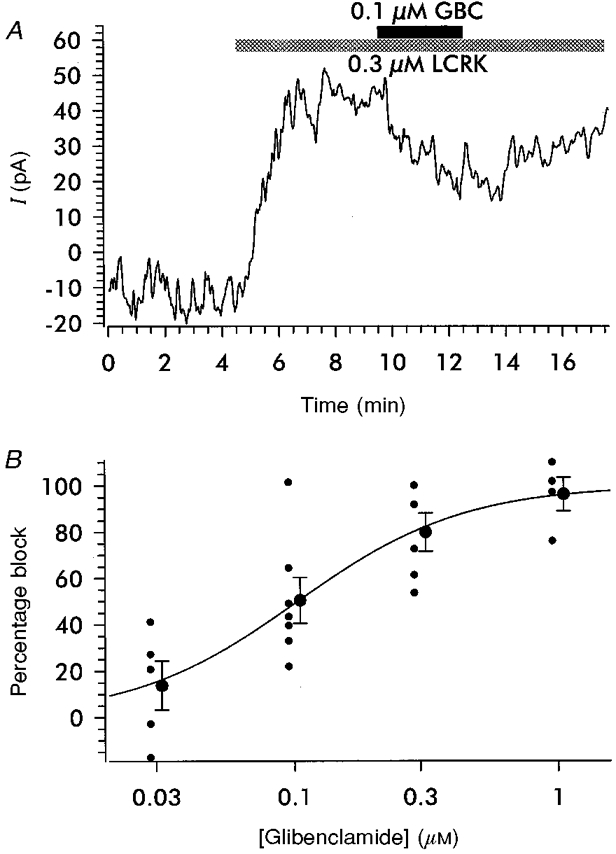
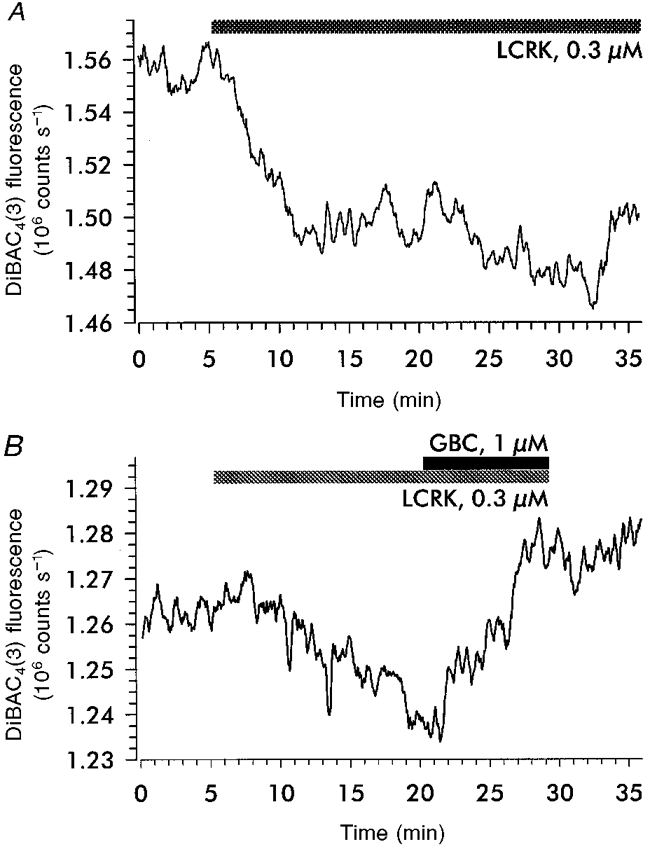
1. Openers of the ATP-sensitive potassium channel (KATP channel) increase and blockers decrease renin secretion. Here we report the effects of levcromakalim (LCRK, a channel opener) and glibenclamide (GBC, a blocker) on membrane potential, whole-cell current and the cytoplasmic Ca2+ concentration of renin-secreting cells (RSC). Studies were performed on afferent arterioles from the kidney of Na+-depleted rats. 2. As monitored with the fluorescent oxonol dye DiBAC4(3), LCRK (0.3 and 1 microM) induced a hyperpolarization of approximately 15 mV which was abolished by GBC (1 microM). 3. Whole-cell current-clamp experiments showed that RSC had a membrane potential of -61 +/- 1 mV (n = 16). LCRK (1 microM) induced a hyperpolarization of 9.9 +/- 0.2 mV (n = 16) which, in the majority of cells, decreased slowly with time. 4. Capacitance measurements showed a strong electrical coupling of the cells in the preparation. 5. At -60 mV, LCRK induced a hyperpolarizing current in a concentration-dependent manner with an EC50 of 152 +/- 31 nM and a maximum current of about 200 pA. 6. Application of GBC (1 microM) produced no effect; however, when applied after LCRK (300 nM), GBC inhibited the opener-induced hyperpolarizing current with an IC50 of 103 +/- 36 nM. 7. LCRK (0.3 and 1 microM) did not significantly affect the cytoplasmic Ca2+ concentration either at rest or after stimulation by angiotensin II. 8. The data show that LCRK induces a GBC-sensitive hyperpolarizing current in rat RSC. This current presumably originates from the activation of KATP channels which pharmacologically resemble those in vascular smooth muscle cells. The stimulatory effect of KATP channel opening on renin secretion is not mediated by a decrease in intracellular Ca2+ concentration.
Figures
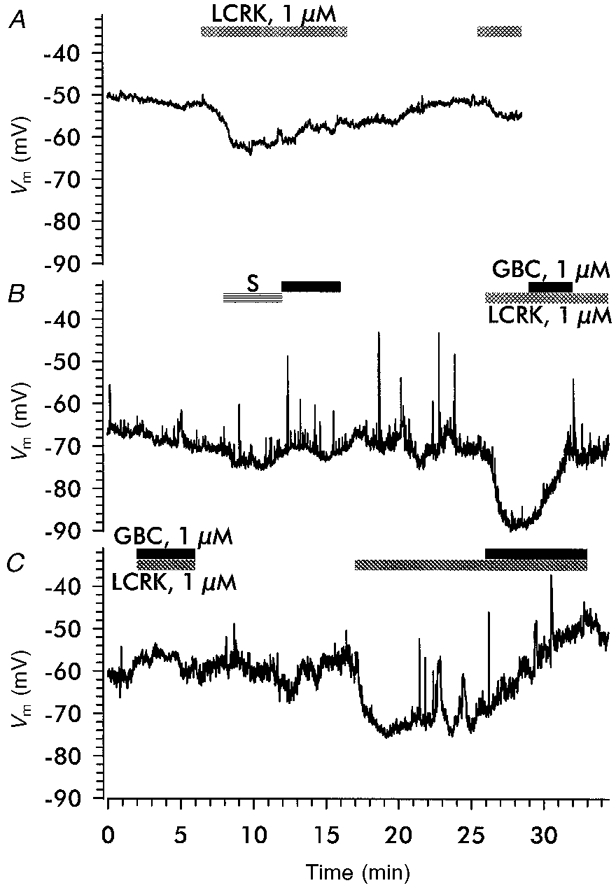
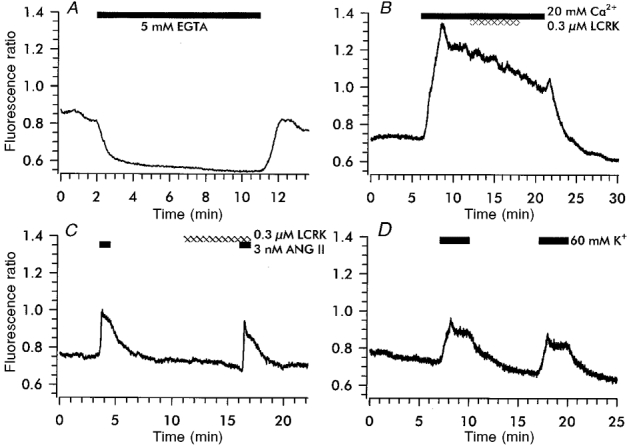
Similar articles
-
Binding and effects of KATP channel openers in the vascular smooth muscle cell line, A10.Br J Pharmacol. 1997 Nov;122(6):1119-26. doi: 10.1038/sj.bjp.0701514. Br J Pharmacol. 1997. PMID: 9401776 Free PMC article.
-
KATP channels are not essential for pressure-dependent control of renin secretion.Pflugers Arch. 1998 Apr;435(5):670-7. doi: 10.1007/s004240050568. Pflugers Arch. 1998. PMID: 9479020
-
ATP-sensitive K+ channel opener acts as a potent Cl- channel inhibitor in vascular smooth muscle cells.J Membr Biol. 1994 Jan;137(1):59-70. doi: 10.1007/BF00234998. J Membr Biol. 1994. PMID: 8006946
-
Pharmacological and molecular characterization of ATP-sensitive K+ channels in the TE671 human medulloblastoma cell line.Eur J Pharmacol. 1999 Apr 9;370(2):179-85. doi: 10.1016/s0014-2999(99)00128-4. Eur J Pharmacol. 1999. PMID: 10323267 Review.
-
Potassium-current oscillation of rat megakaryocytes: As a model system for drug evaluation (Review).Int J Mol Med. 1999 Aug;4(2):163-9. doi: 10.3892/ijmm.4.2.163. Int J Mol Med. 1999. PMID: 10402483 Review.
Cited by
-
The influence of extracellular and intracellular calcium on the secretion of renin.Pflugers Arch. 2013 Jan;465(1):59-69. doi: 10.1007/s00424-012-1107-x. Epub 2012 Apr 28. Pflugers Arch. 2013. PMID: 22538344 Free PMC article. Review.
-
ATP-sensitive potassium channels induced in liver cells after transfection with insulin cDNA and the GLUT 2 transporter regulate glucose-stimulated insulin secretion.FASEB J. 2003 Sep;17(12):1682-4. doi: 10.1096/fj.02-0051fje. Epub 2003 Jul 18. FASEB J. 2003. PMID: 12958175 Free PMC article.
-
Electrophysiological and molecular characterization of the inward rectifier in juxtaglomerular cells from rat kidney.J Physiol. 2004 Oct 15;560(Pt 2):365-76. doi: 10.1113/jphysiol.2004.070359. Epub 2004 Jul 29. J Physiol. 2004. PMID: 15284349 Free PMC article.
-
Connexin Hemichannels Contribute to the Activation of cAMP Signaling Pathway and Renin Production.Int J Mol Sci. 2020 Jun 23;21(12):4462. doi: 10.3390/ijms21124462. Int J Mol Sci. 2020. PMID: 32585970 Free PMC article.
-
Effect of adenosine on membrane potential and Ca2+ in juxtaglomerular cells. Comparison with angiotensin II.Kidney Blood Press Res. 2008;31(2):94-103. doi: 10.1159/000119712. Epub 2008 Mar 6. Kidney Blood Press Res. 2008. PMID: 18322364 Free PMC article.
References
-
- Ashcroft SJH, Ashcroft FM. Properties and functions of ATP-sensitive K-channels. Cellular Signalling. 1990;2:197–214. 10.1016/0898-6568(90)90048-F. - DOI - PubMed
-
- Bräuner T, Hülser DF, Strasser RJ. Comparative measurements of membrane potentials with microelectrodes and voltage-sensitive dyes. Biochimica et Biophysica Acta. 1984;771:208–216. - PubMed
-
- Bray KM, Quast U. A specific binding site for K+ channel openers in rat aorta. Journal of Biological Chemistry. 1992;267:11689–11692. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous