

BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth
- PMID: 10359578
- PMCID: PMC2193079
- DOI: 10.1084/jem.189.11.1747
BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth
Abstract
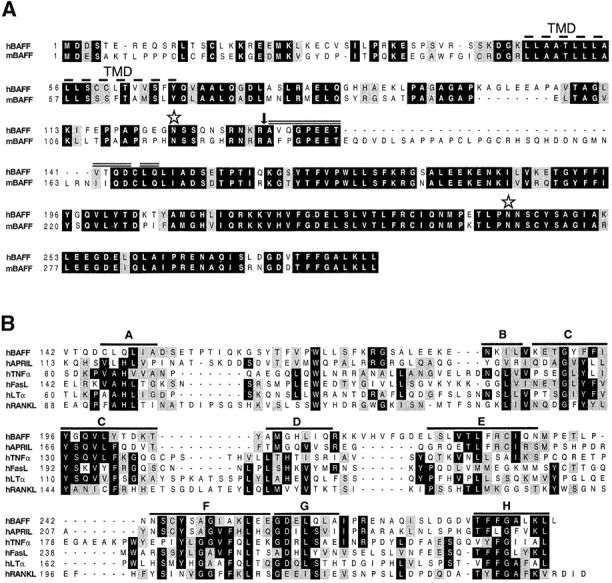
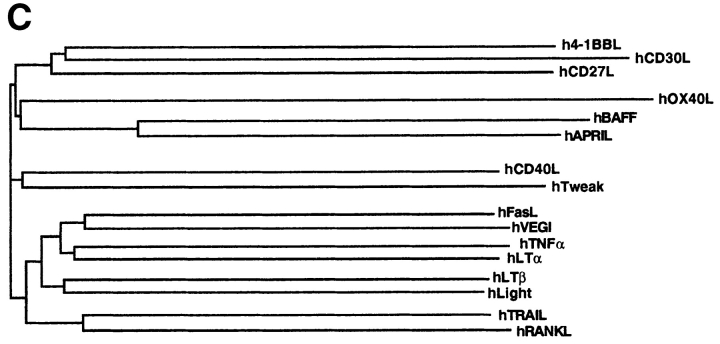
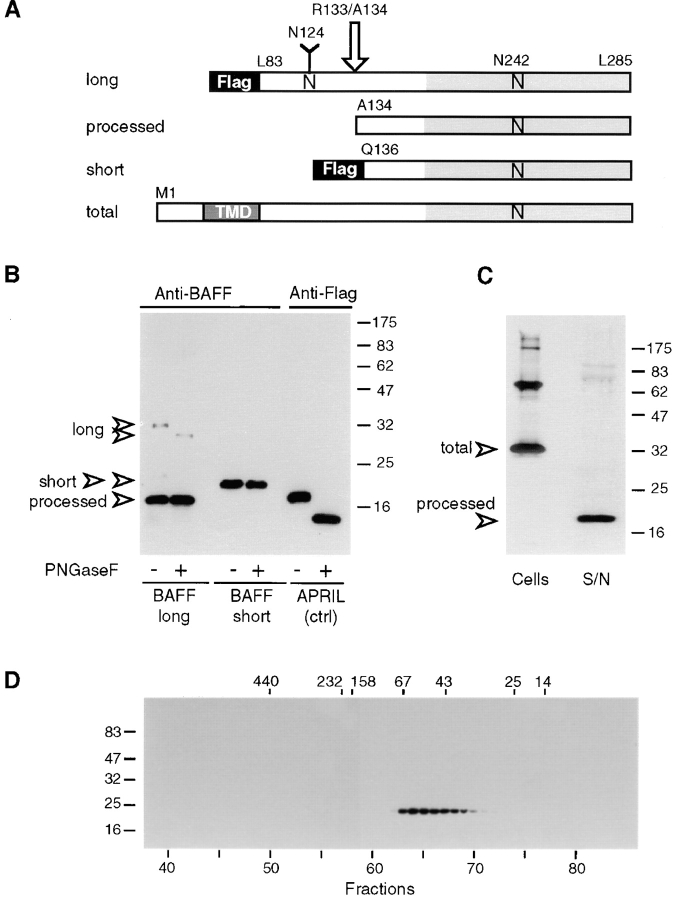
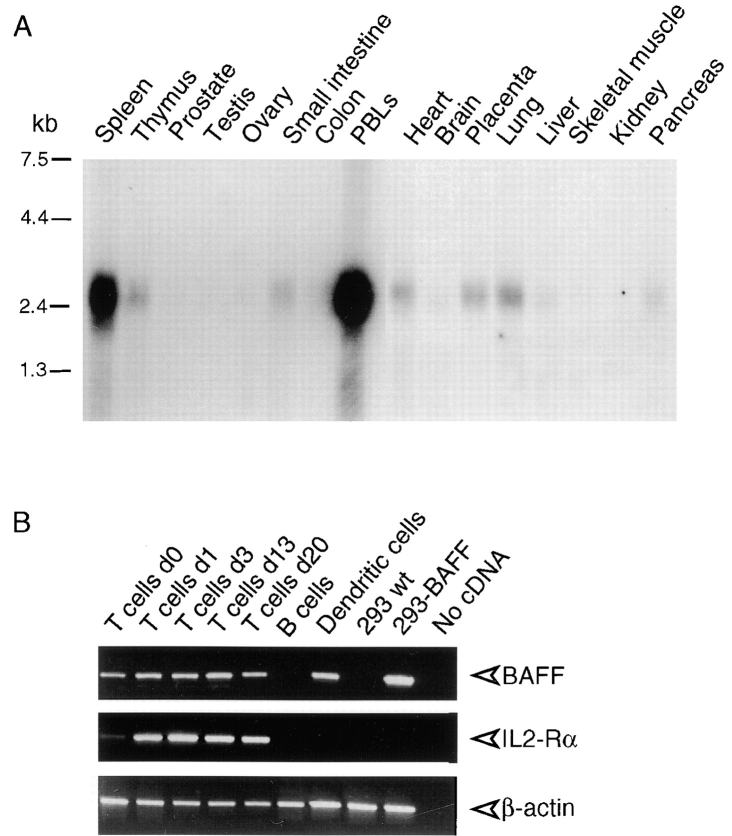
Members of the tumor necrosis factor (TNF) family induce pleiotropic biological responses, including cell growth, differentiation, and even death. Here we describe a novel member of the TNF family, designated BAFF (for B cell activating factor belonging to the TNF family), which is expressed by T cells and dendritic cells. Human BAFF was mapped to chromosome 13q32-34. Membrane-bound BAFF was processed and secreted through the action of a protease whose specificity matches that of the furin family of proprotein convertases. The expression of BAFF receptor appeared to be restricted to B cells. Both membrane-bound and soluble BAFF induced proliferation of anti-immunoglobulin M-stimulated peripheral blood B lymphocytes. Moreover, increased amounts of immunoglobulins were found in supernatants of germinal center-like B cells costimulated with BAFF. These results suggest that BAFF plays an important role as costimulator of B cell proliferation and function.
Figures
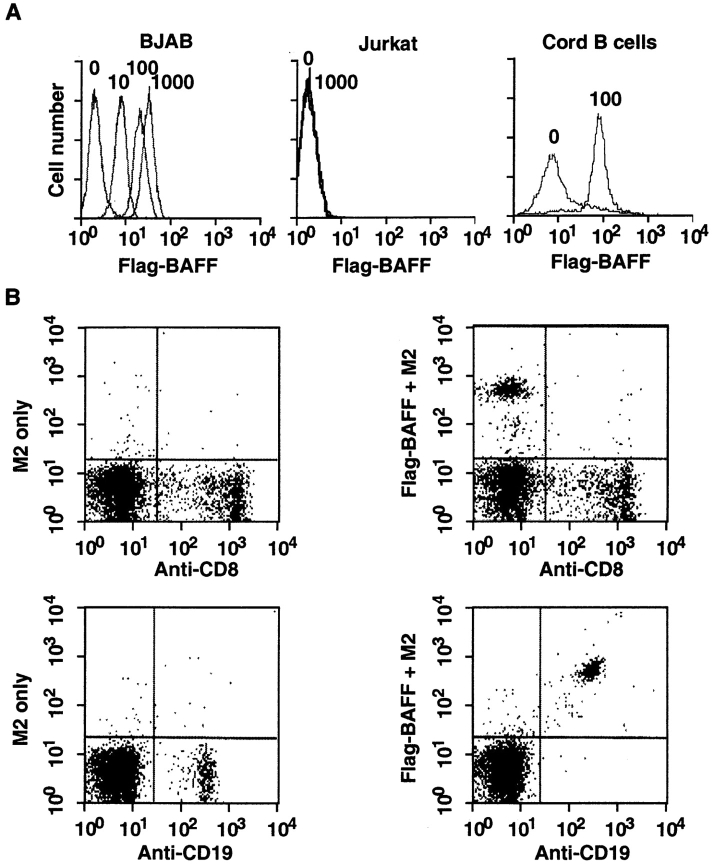
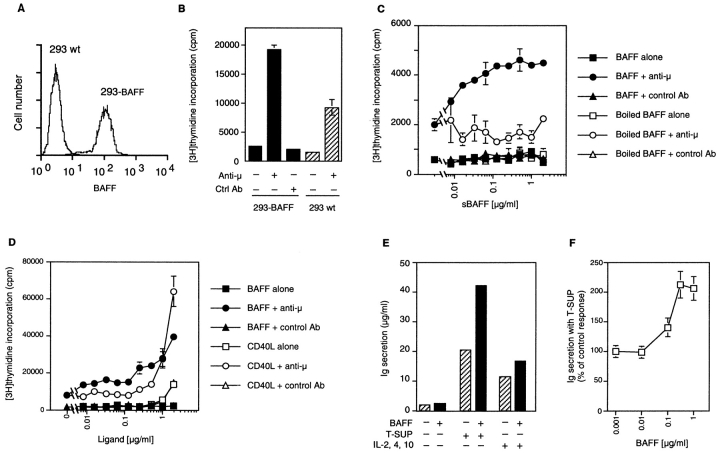
References
-
- Smith CA, Farrah T, Goodwin RG. The TNF-receptor superfamily of cellular and viral proteins: activation, costimulation, and death. Cell. 1994;76:959–962. - PubMed
-
- Vassalli P. The pathophysiology of tumor necrosis factors. Annu Rev Immunol. 1992;10:411–452. - PubMed
-
- De Togni P, Goellner J, Ruddle NH, Streeter PR, Fick A, Mariathasan S, Smith SC, Carlson R, Shornick LP, Strauss-Schoenberger J, et al. Abnormal development of peripheral lymphoid organs in mice deficient in lymphotoxin. Science. 1994;264:703–707. - PubMed
-
- Koni PA, Sacca R, Lawton P, Browning JL, Ruddle NH, Flavell RA. Distinct roles in lymphoid organogenesis for lymphotoxins alpha and beta revealed in lymphotoxin beta-deficient mice. Immunity. 1997;6:491–500. - PubMed
-
- Amakawa R, Hakem A, Kundig TM, Matsuyama T, Simard JJ, Timms E, Wakeham A, Mittruecker HW, Griesser H, Takimoto H, et al. Impaired negative selection of T cells in Hodgkin's disease antigen CD30-deficient mice. Cell. 1996;84:551–562. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
