

doi: 10.1091/mbc.10.6.1923.
Erp1p and Erp2p, partners for Emp24p and Erv25p in a yeast p24 complex
Affiliations
- PMID: 10359606
- PMCID: PMC25390
- DOI: 10.1091/mbc.10.6.1923
Item in Clipboard
Erp1p and Erp2p, partners for Emp24p and Erv25p in a yeast p24 complex
Mol Biol Cell.
1999 Jun.
Free PMC article
Abstract
Six new members of the yeast p24 family have been identified and characterized. These six genes, named ERP1-ERP6 (for Emp24p- and Erv25p-related proteins) are not essential, but deletion of ERP1 or ERP2 causes defects in the transport of Gas1p, in the retention of BiP, and deletion of ERP1 results in the suppression of a temperature-sensitive mutation in SEC13 encoding a COPII vesicle coat protein. These phenotypes are similar to those caused by deletion of EMP24 or ERV25, two previously identified genes that encode related p24 proteins. Genetic and biochemical studies demonstrate that Erp1p and Erp2p function in a heteromeric complex with Emp24p and Erv25p.
Figures
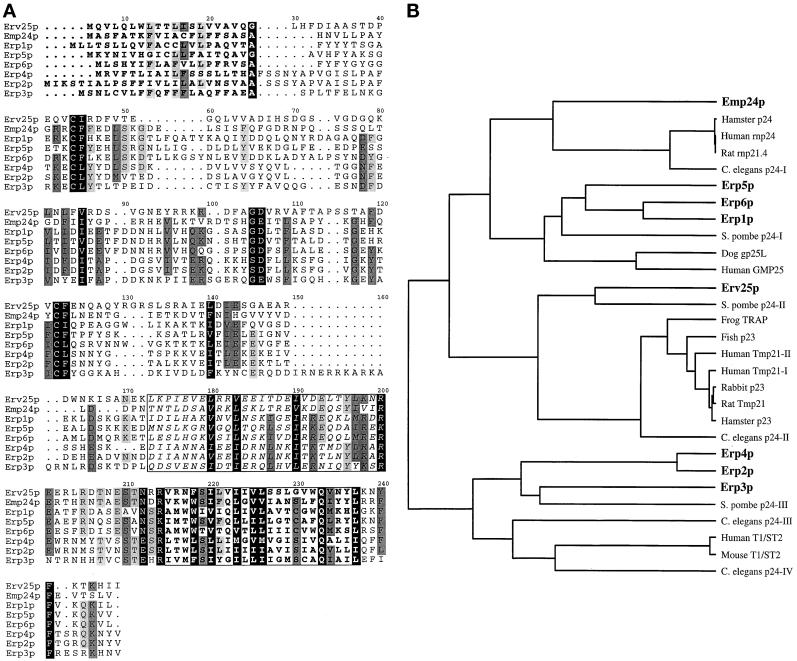
Yeast p24 proteins and the relationship of currently identified p24 genes. (A) Alignment of the eight p24 members in S. cerevisiae. The predicted N-terminal signal sequences are marked in bold. The putative coiled coil domains are boxed, and in italics and the predicted transmembrane region is boxed and in bold. All yeast p24 proteins, as in other species, contain two conserved cysteine residues within the N-terminal region of the protein that is likely to be in the lumen of a secretory compartment or vesicle. Each yeast p24 protein contains a conserved glutamine residue within the transmembrane domain and a conserved phenylalanine residue (implicated in COPII binding; Dominguez et al. 1998) in the membrane-proximal region of the short cytoplasmic C-terminus. Yeast α family p24 proteins (Erp1p, Erp5p, and Erp6p) contain close matches to the COPI binding motifs KKXX and KXKXX (Cosson and Letourneur, 1994). All the yeast α and β family members (Erp1p, Erp5p, Erp6p, and Emp24p) contain a polar glutamine residue within the transmembrane domain (position 222). This residue in Chop24 has been implicated in ER localization (Fiedler and Rothman, 1997). (B) Relationship between predicted p24 protein sequences currently deposited in public domain databases. The phylogenetic tree shown was generated using the UPMGA clustering method of NEIGHBOR from the PHYLIP package (Felsenstein, 1989), the distance matrix being calculated using PROTDIST.
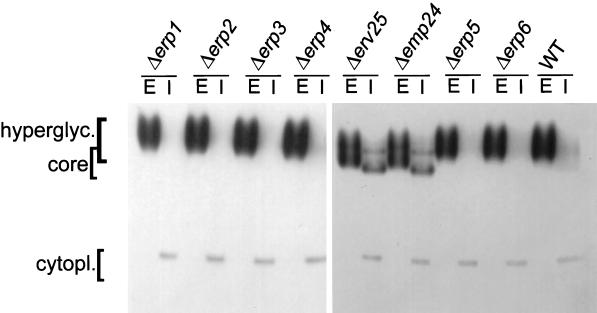
Invertase secretion in wild-type and p24 mutant cells. Exponentially dividing cells were transferred to low-glucose media for 1 h to induce invertase synthesis. Spheroplasts were prepared and pelleted by centrifugation to obtain the cytoplasmic (I) and periplasmic (E) fractions. Invertase was detected by an in-gel activity assay. The cytoplasmic, core, and hyperglycosylated forms of invertase are indicated.
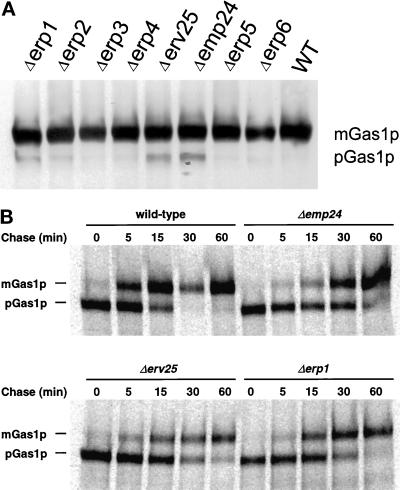
Maturation of Gas1p in wild-type and p24 deletion strains. (A) Whole-cell extracts were prepared from exponentially growing cells, separated by SDS-PAGE, and detected by Western blotting using an anti-Gas1p antibody. Migration of mature (mGas1p) and precursor (pGas1p) forms of Gas1p are indicated. (B) Extracts were prepared from wild-type, Δemp24, Δerv25, and Δerp1 cells after a 5-min pulse with 35S-labeled amino acids followed by chase for the indicated times. Gas1p was precipitated from the extracts, and immune complexes were precipitated, separated by SDS-PAGE, and detected by autoradiography. Migration of mature (mGas1p) and precursor (pGas1p) forms of Gas1p is indicated.
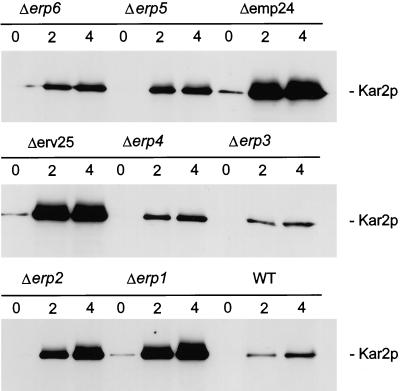
Analysis of Kar2p secretion in p24 mutant strains. Exponentially dividing cells were washed and resuspended in fresh medium. Proteins of the culture medium of 0.3 OD600 unit equivalents were TCA precipitated at the indicated times and resolved by SDS-PAGE. Kar2p was detected by Western blotting with anti-Kar2p antiserum.
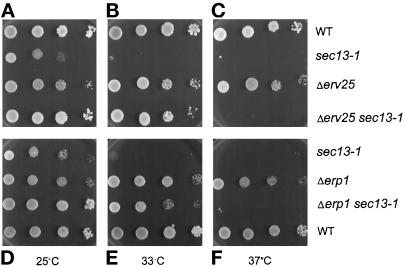
Suppression of sec13-1 by p24 deletion mutations. Each p24 deletion mutant was crossed with a sec13-1 temperature-sensitive mutant, and segregants from a tetratype tetrad were tested for growth at different temperatures on YEPD medium. Results shown are at 25°C, a permissive sec13-1 temperature (A and D), 33°C (B and E), and 37°C (C and F), nonpermissive temperatures for sec13-1 mutants.
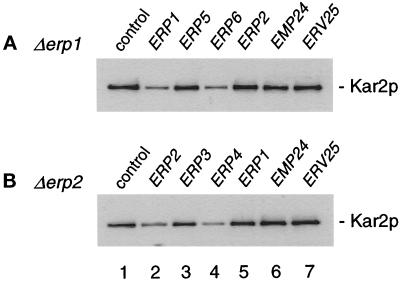
Genetic redundancy among p24 genes. Individual p24 genes were overexpressed from the TPI promotor in erp1 or erp2 mutant cells grown at 30°C in YPD. Exponentially dividing cells were washed and resuspended in fresh medium. Proteins of the culture medium of 0.3 OD600 unit equivalents were TCA precipitated after 4.5 h and resolved by SDS-PAGE. Kar2p was detected by Western blotting with anti-Kar2p antiserum.
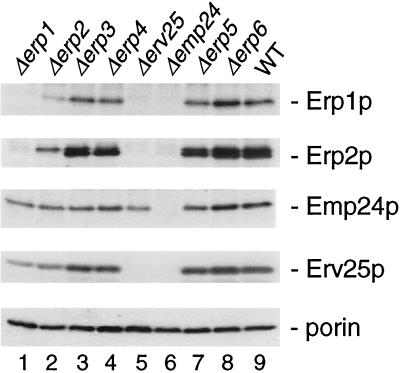
Emp24p, Erv25p, Erp1p, and Erp2p protein levels are interdependent. Total cell extracts from wild-type and p24 mutant cells were separated by SDS-PAGE, and proteins were detected on Western blots using Emp24p, Erv25p, Erp1p, or Erp2p antisera as indicated. A mAb to porin, a mitochondrial membrane protein unrelated to this study, was used as a loading control.
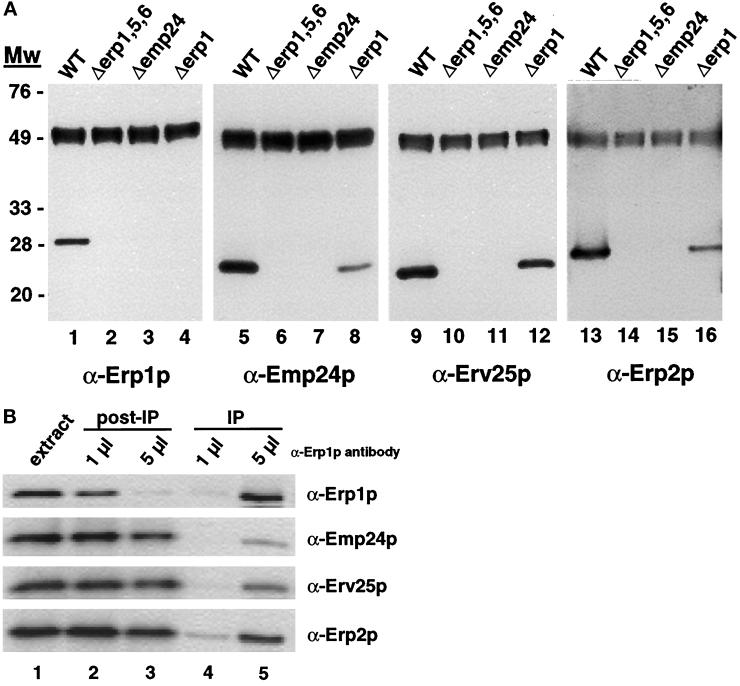
Immunoprecipitation of Erp1p from wild-type and p24 mutant strains. (A) Erp1p was precipitated from solubilized extracts as described in MATERIALS AND METHODS. Precipitates were resolved on 12% SDS-polyacrylamide gels, transferred to nitrocellulose, and probed with antibodies against Erp1p (lanes 1–4), Emp24p (lanes 5–8), Erv25p (lanes 9–12), and Erp2p (lanes 13–16). The immunoreactive signal detected at ∼48 kDa corresponds to the heavy chain of the anti-Erp1p antibodies used for the precipitation. (B) Erp1p was quantitatively immunoprecipitated from solubilized extracts prepared from wild-type cells as described in MATERIALS AND METHODS. Equivalent proportions of the extract before (lane 1) and after (lanes 2 and 3) immunoprecipitation and of the immunoprecipitate (lanes 4 and 5) were resolved on 12% SDS-polyacrylamide gels, transferred to nitrocellulose, and probed with antibodies against Erp1p, Erp2p, Emp24p, and Erv25p. Immunoprecipitations were performed with either 1 μl (lanes 2 and 4) or 5 μl (lanes 3 and 5) of Erp1p antiserum.
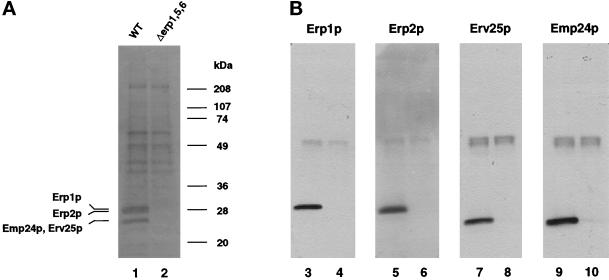
Immunoprecipitation of Erp1p from radiolabeled wild-type and erp1/5/6 mutant cells. Erp1p was precipitated from solubilized extracts prepared from radiolabeled wild-type or erp1/5/6 mutant cells (A) or unlabeled cells (B) as described in MATERIALS AND METHODS. Immunoprecipitates were resolved on 12% SDS-polyacrylamide gels and either dried and exposed to film (A) or transferred to nitrocellulose (B) and probed with antibodies against Erp1p (lanes 3 and 4), Erp2p (lanes 5 and 6), Erv25p (lanes 7 and 8), and Emp24p (lanes 9 and 10). Immunoprecipitates in lanes 1, 3, 5, 7, and 9 were prepared from wild-type cells; those in lanes 2, 4, 6, 8, and 10 were prepared from erp1/5/6 mutant cells. The immunoreactive protein detected at ∼48 kDa corresponds to the heavy chain of the anti-Erp1p antibodies used for the immunoprecipitation.
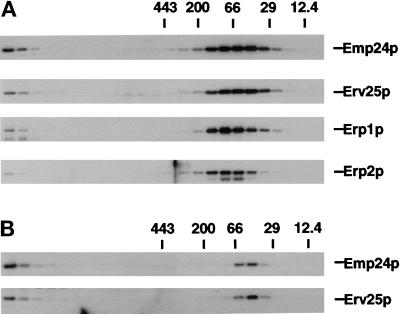
Gel filtration chromatography. Wild-type (A) and erp1/erp5/erp6 mutant (B) yeast cells were converted to spheroplasts and lysed, and the cleared extract was applied to a Superose 6 HR 10/30 gel filtration column. The fractionation was calibrated by parallel runs of molecular weight markers. Individual p24 proteins were detected by Western blot analysis.
References
-
- Barlowe C, Orci L, Yeung T, Hosobuchi M, Hamamoto S, Salama N, Rexach MF, Ravazzola M, Amherdt M, Schekman R. COPII: a membrane coat formed by Sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell. 1994;77:895–907. - PubMed
-
- Bednarek SY, Ravazzola M, Hosobuchi M, Amherdt M, Perrelet A, Schekman R, Orci L. COPI- and COPII-coated vesicles bud directly from the endoplasmic reticulum in yeast. Cell. 1995;83:1183–1196. - PubMed
-
- Belden WJ, Barlowe C. Erv25p, a component of COPII-coated vesicles, forms a complex with Emp24p that is required for efficient endoplasmic reticulum to Golgi transport. J Biol Chem. 1996;271:26939–26946. - PubMed
-
- Blum R, Feick P, Puype M, Vandekerckhove J, Klengel R, Nastainczyk W, Schulz I. Tmp21 and p24a, two type I proteins enriched in pancreatic microsomal membranes, are members of a protein family involved in vesicular trafficking. J Biol Chem. 1996;271:17183–17189. - PubMed
-
- Cosson P, Letourneur F. Coatomer interaction with di-lysine endoplasmic reticulum retention motifs. Science. 1994;263:1629–1631. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
