

Activation of utrophin promoter by heregulin via the ets-related transcription factor complex GA-binding protein alpha/beta
- PMID: 10359616
- PMCID: PMC25417
- DOI: 10.1091/mbc.10.6.2075
Activation of utrophin promoter by heregulin via the ets-related transcription factor complex GA-binding protein alpha/beta
Abstract
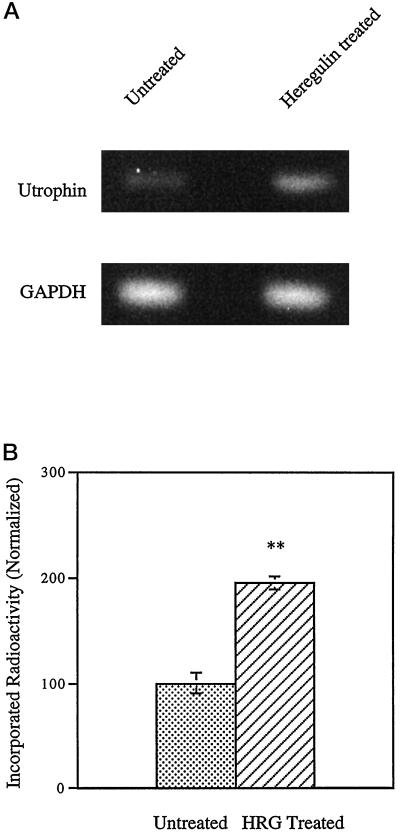
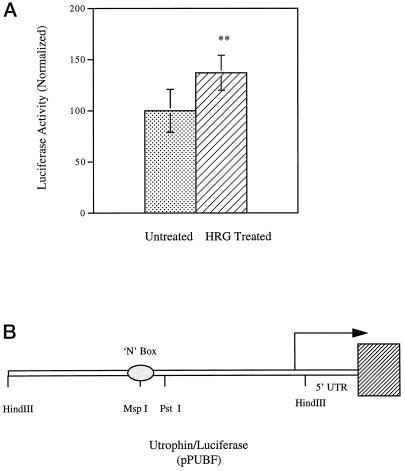
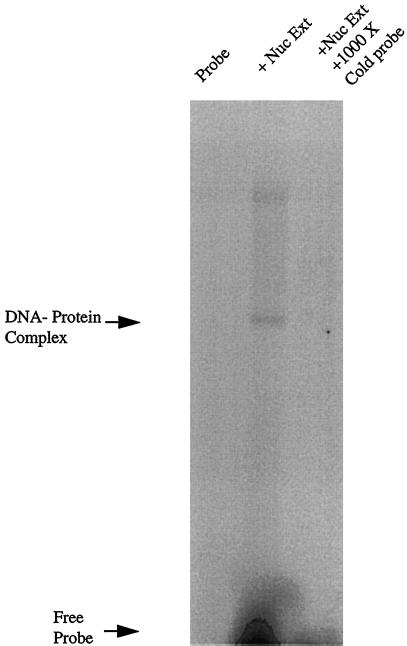
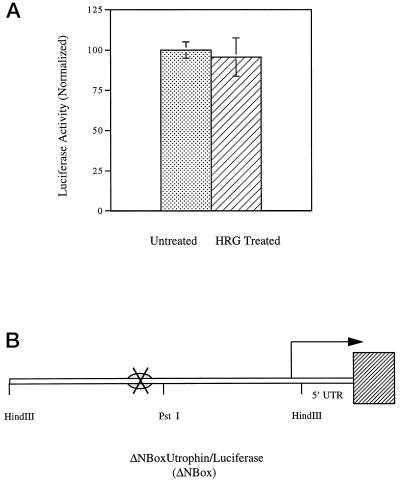
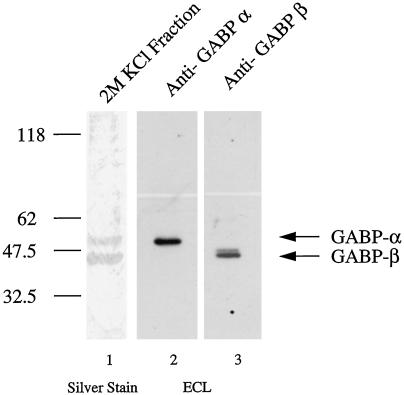
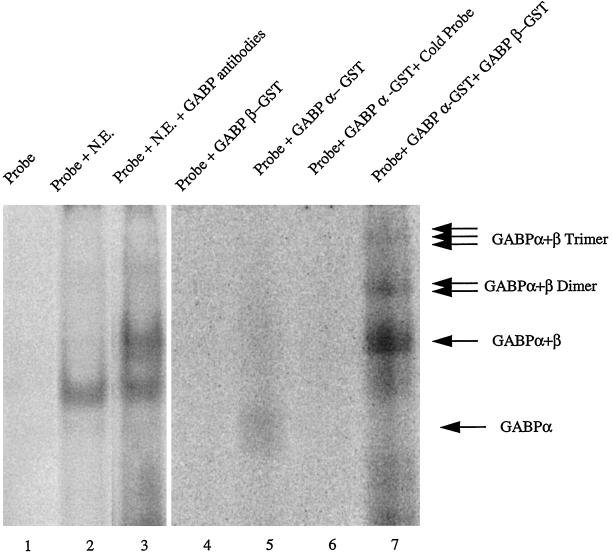
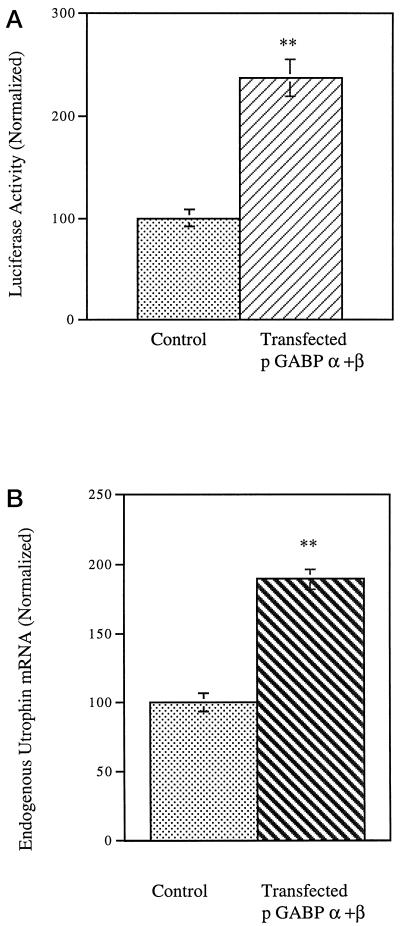
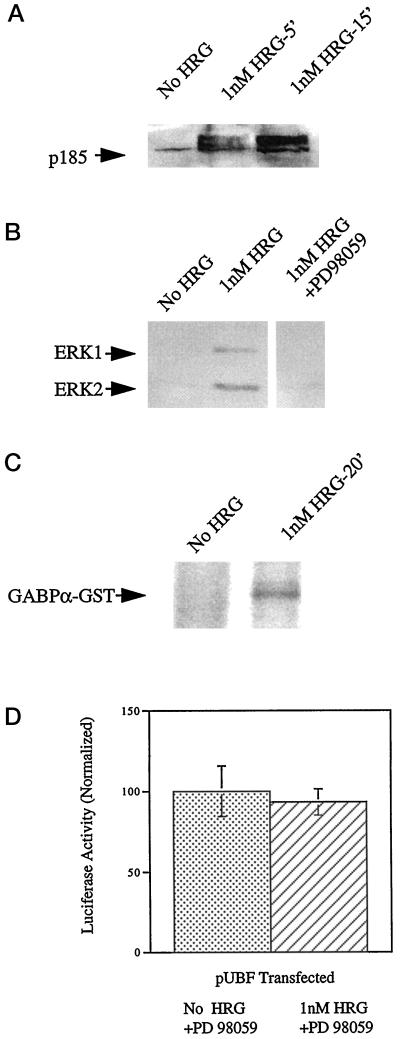
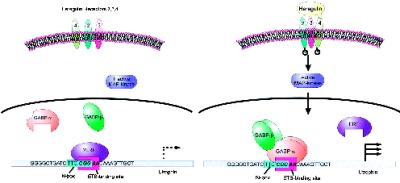
Utrophin/dystrophin-related protein is the autosomal homologue of the chromosome X-encoded dystrophin protein. In adult skeletal muscle, utrophin is highly enriched at the neuromuscular junction. However, the molecular mechanisms underlying regulation of utrophin gene expression are yet to be defined. Here we demonstrate that the growth factor heregulin increases de novo utrophin transcription in muscle cell cultures. Using mutant reporter constructs of the utrophin promoter, we define the N-box region of the promoter as critical for heregulin-mediated activation. Using this region of the utrophin promoter for DNA affinity purification, immunoblots, in vitro kinase assays, electrophoretic mobility shift assays, and in vitro expression in cultured muscle cells, we demonstrate that ets-related GA-binding protein alpha/beta transcription factors are activators of the utrophin promoter. Taken together, these results suggest that the GA-binding protein alpha/beta complex of transcription factors binds and activates the utrophin promoter in response to heregulin-activated extracellular signal-regulated kinase in muscle cell cultures. These findings suggest methods for achieving utrophin up-regulation in Duchenne's muscular dystrophy as well as mechanisms by which neurite-derived growth factors such as heregulin may influence the regulation of utrophin gene expression and subsequent enrichment at the neuromuscular junction of skeletal muscle.
Figures
References
-
- Ausubel F, et al. Current Protocols in Molecular Biology. New York: Wiley; 1995.
-
- Brown RH. Dystrophin-associated proteins and muscular dystrophies. Annu Rev Med. 1997;48:457–466. - PubMed
-
- Brown TA, McKnight SL. Specificities of protein-protein and protein-DNA interaction of GABP alpha and two newly defined ets-related proteins. Genes & Dev. 1992;6:2502–2512. - PubMed
-
- Campbell KP. Three muscular dystrophies: loss of cytoskeletal extracellular matrix linkage. Cell. 1996;80:675–679. - PubMed
-
- Campbell KP, Crosbie RH. Muscular dystrophy. Utrophin to the rescue. Nature. 1996;384:308–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
