

A nonhuman primate model for the selective elimination of CD8+ lymphocytes using a mouse-human chimeric monoclonal antibody
- PMID: 10362819
- PMCID: PMC1866630
- DOI: 10.1016/S0002-9440(10)65450-8
A nonhuman primate model for the selective elimination of CD8+ lymphocytes using a mouse-human chimeric monoclonal antibody
Abstract
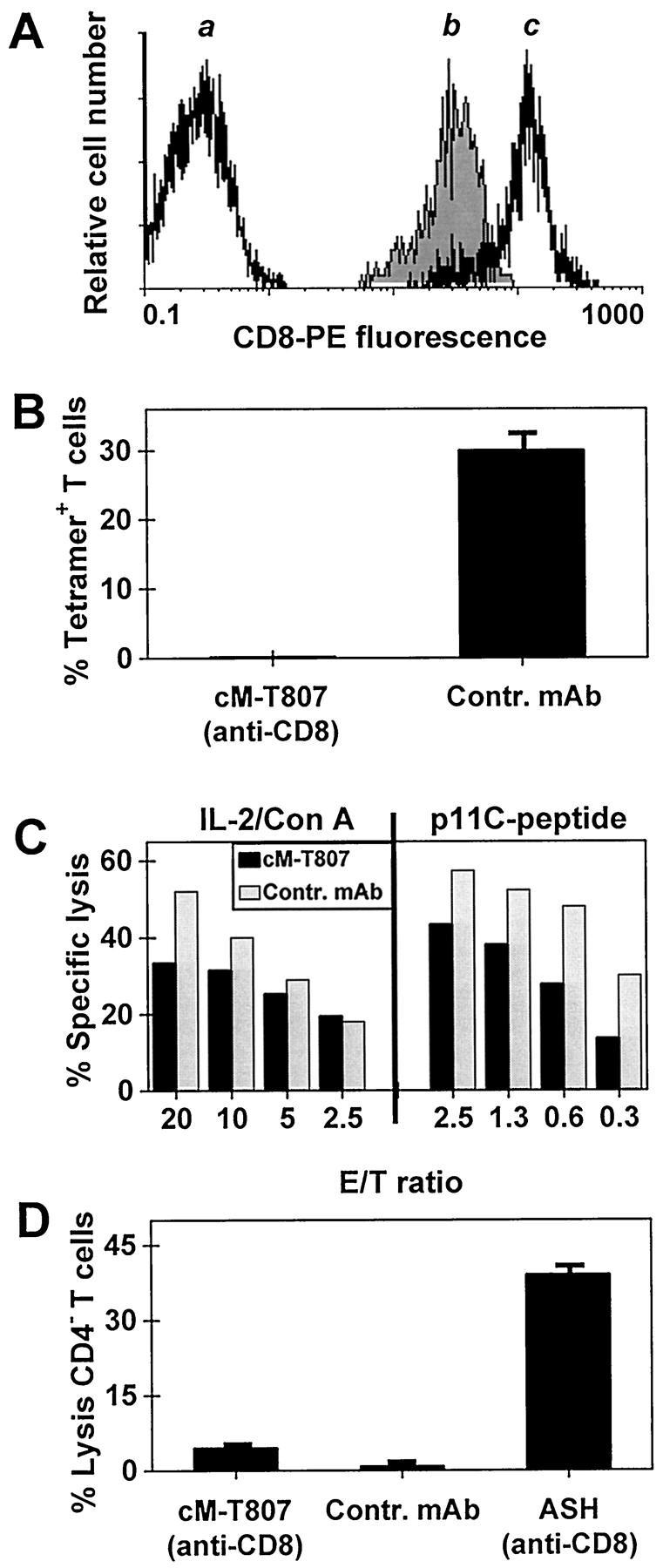
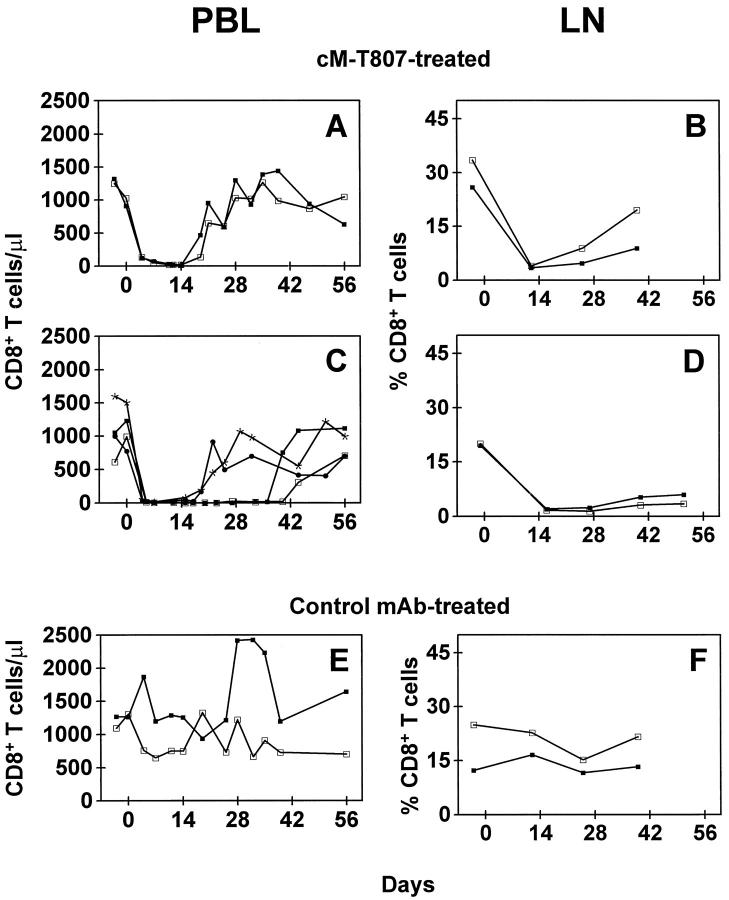
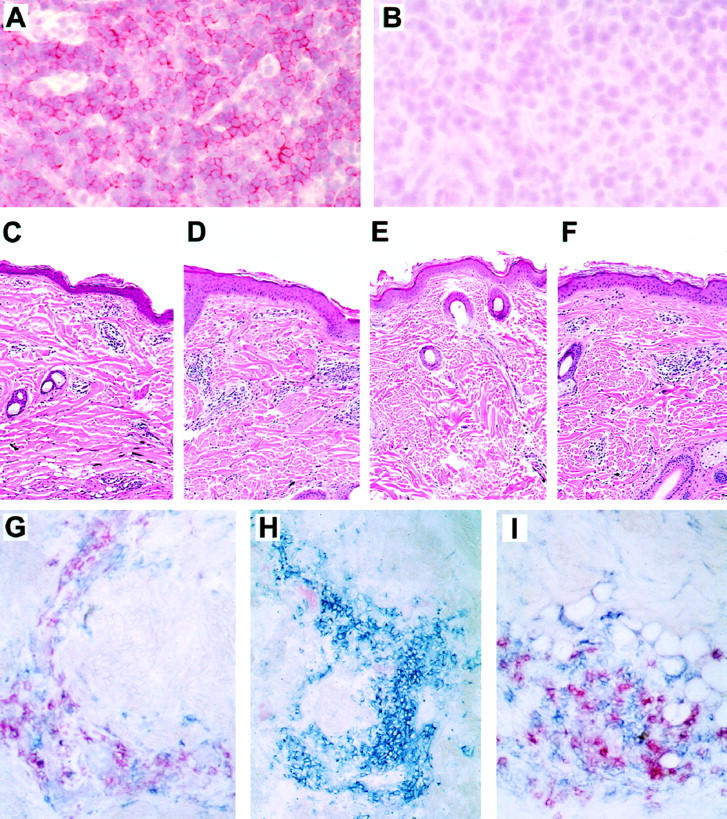
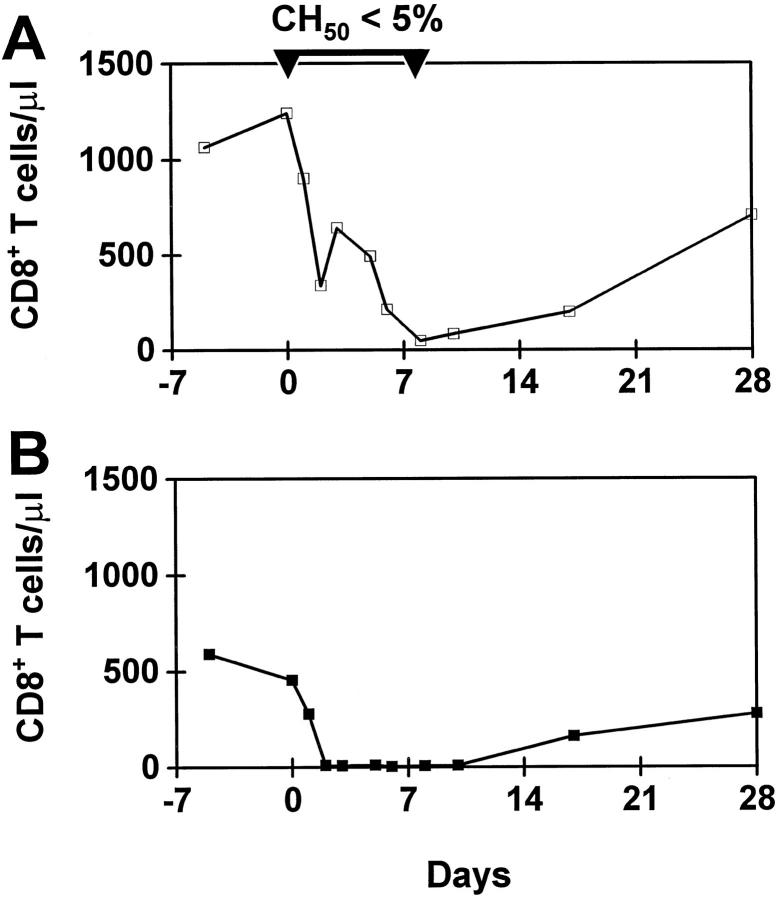
Nonhuman primates provide valuable animal models for human diseases. However, studies assessing the role of cell-mediated immune responses have been difficult to perform in nonhuman primates. We have shown that CD8+ lymphocyte-mediated immunity in rhesus monkeys can be selectively eliminated using the mouse-human chimeric anti-CD8 monoclonal antibody cM-T807. In vitro, this antibody completely blocked antigen-specific expansion of cytotoxic T cells and decreased major histocompatibility complex class I-restricted, antigen-specific lysis of target cells but did not mediate complement-dependent cell lysis. In vivo administration of cM-T807 in rhesus monkeys resulted in near total depletion of CD8+ T cells from the blood and lymph nodes for up to 6 weeks. This depletion was not solely complement-dependent and persisted longer in adults than in juveniles. Preservation of B cell and CD4+ T cell function in monkeys depleted of CD8+ lymphocytes was demonstrated by their ability to develop humoral immune responses to the administered chimeric monoclonal antibody. Furthermore, during CD8+ lymphocyte depletion, monkeys developed delayed-type hypersensitivity reactions comprised only of CD4+ T cells but not CD8+ T cells. This CD8+ lymphocyte depletion model should prove useful in defining the role of cell-mediated immune responses in controlling infectious diseases in nonhuman primates.
Figures
References
-
- Yap KL, Ada GL, McKenzie IFC: Transfer of specific cytotoxic T lymphocytes protects mice inoculated with influenza virus. Nature 1978, 273:238-239 - PubMed
-
- Riddell SR, Greenberg PD: Principles for adoptive T cell therapy of human viral diseases. Annu Rev Immunol 1995, 13:545-586 - PubMed
-
- Fung-Leung W-P, Schilham MW, Rahemtulla A, Kundig TM, Vollenweider M, Potter J, van Ewijk W, Mak TW: CD8 is needed for development of cytotoxic T cells but not helper T cells. Cell 1991, 65:443-449 - PubMed
-
- Nesic D, Vukmanovic S: MHC class I is required for peripheral accumulation of CD8+ thymic emigrants. J Immunol 1998, 160:3705-3712 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
