

The Drosophila p38 MAPK pathway is required during oogenesis for egg asymmetric development
- PMID: 10364162
- PMCID: PMC316763
- DOI: 10.1101/gad.13.11.1464
The Drosophila p38 MAPK pathway is required during oogenesis for egg asymmetric development
Abstract
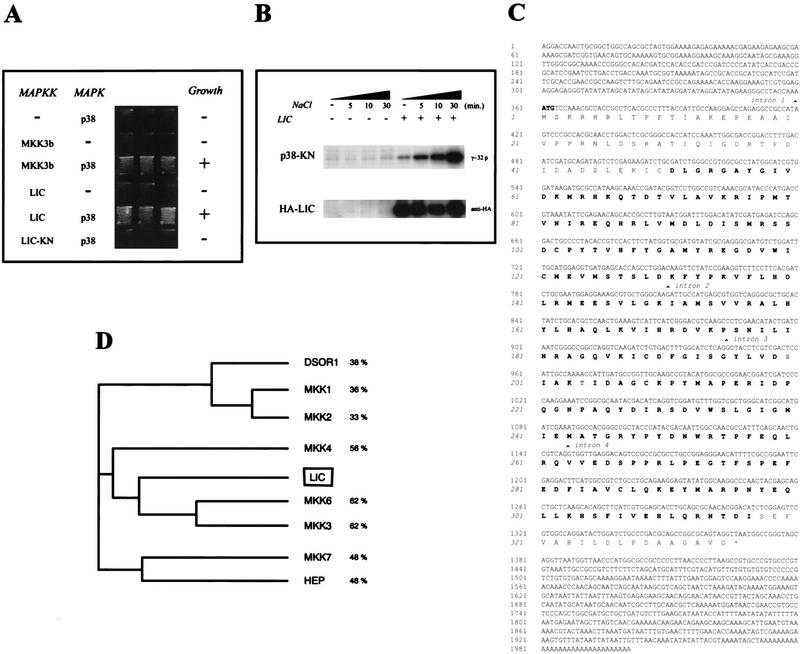
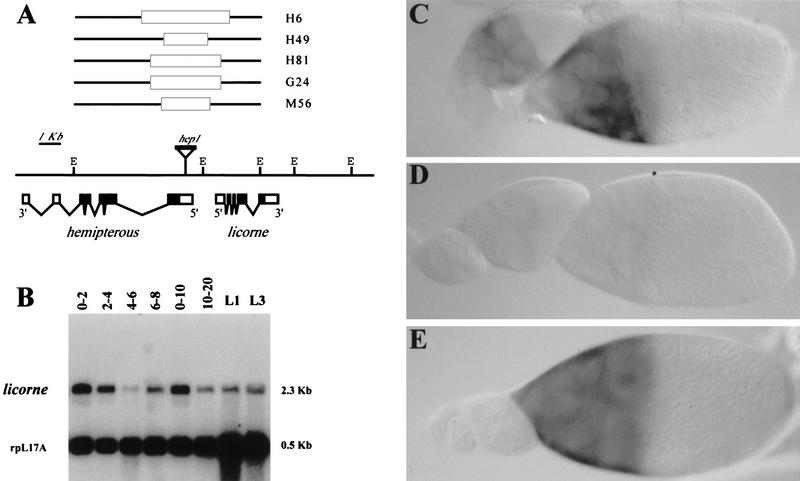
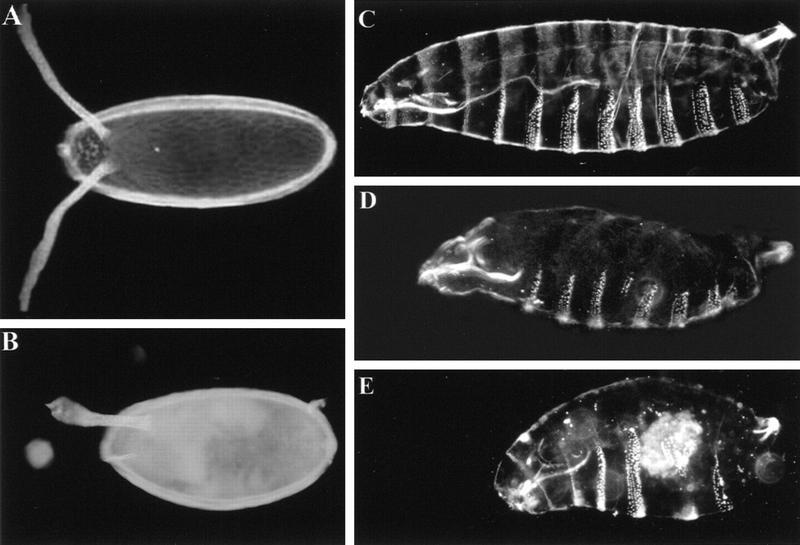
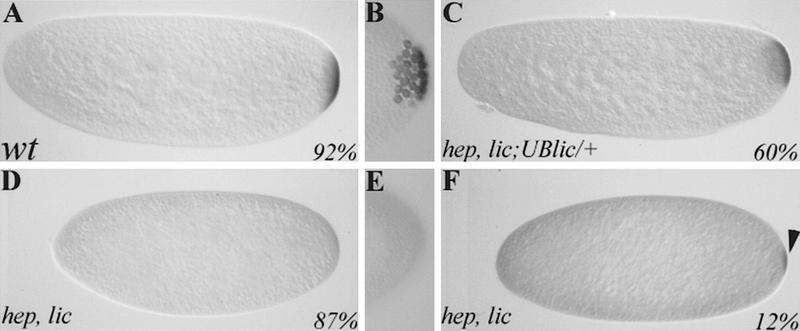
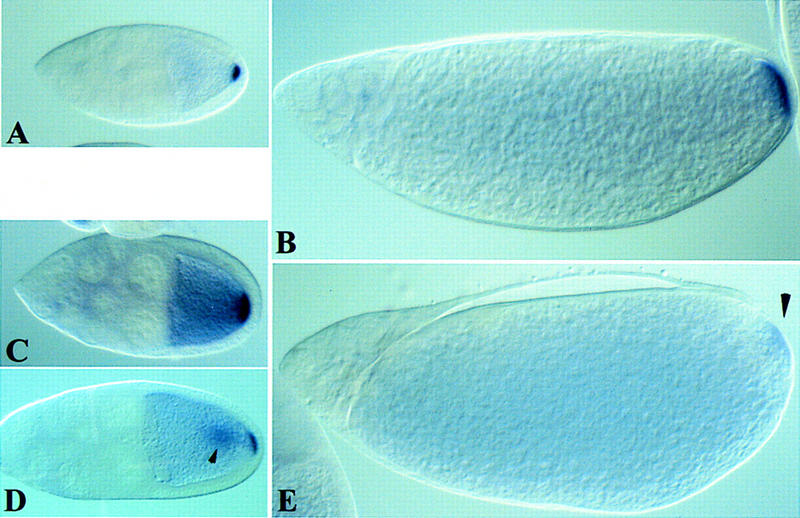
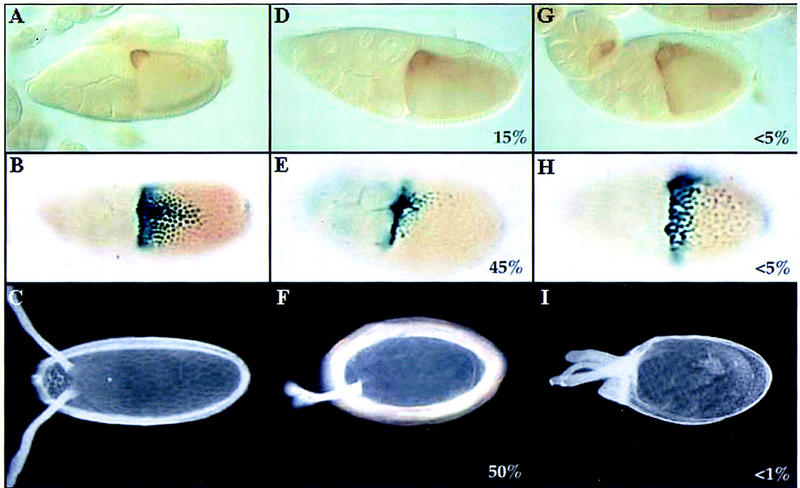
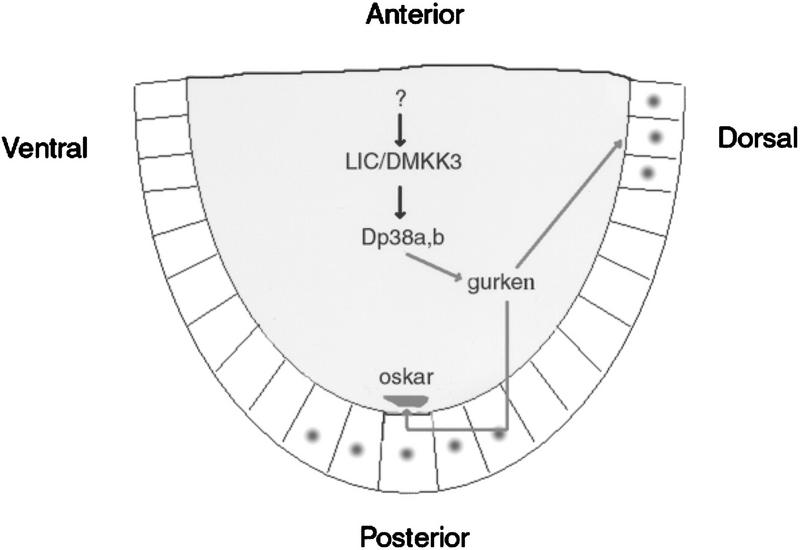
In mammalian cells, the p38 mitogen-activated protein kinase (MAPK) pathway is activated in response to a variety of environmental stresses and inflammatory stimuli. However, the role of p38 MAPK signaling in unchallenged conditions remains largely unknown. We have isolated mutations in a Drosophila p38 MAPKK gene homolog, licorne (lic), and show that during oogenesis, lic is required in the germ line for correct asymmetric development of the egg. In lic mutant egg chambers, oskar mRNA posterior localization is not properly maintained, resulting in anteroposterior patterning defects in the embryo. Furthermore, lic loss-of-function in the germ line leads to reduced EGF receptor activity in dorsal follicle cells and ventralization of the egg shell. Both these defects are associated with a diminution of gurken protein levels in the oocyte. Our phenotypic data argue for a role of lic in a post-transcriptional regulation of the grk gene. Furthermore, they show that in addition to the well-characterized Ras/Raf/ERK MAPK pathway acting in the follicle cells, another related signaling cascade, the p38 MAPK pathway, is required in the germ line for correct axes determination. These results provide the first genetic demonstration of an essential function for a p38 pathway during development.
Figures
Similar articles
-
Post-transcriptional regulation of gurken by encore is required for axis determination in Drosophila.Development. 1997 Dec;124(23):4801-10. doi: 10.1242/dev.124.23.4801. Development. 1997. PMID: 9428416
-
The mago nashi gene is required for the polarisation of the oocyte and the formation of perpendicular axes in Drosophila.Curr Biol. 1997 Jul 1;7(7):468-78. doi: 10.1016/s0960-9822(06)00218-1. Curr Biol. 1997. PMID: 9210377
-
Drosophila rhino encodes a female-specific chromo-domain protein that affects chromosome structure and egg polarity.Genetics. 2001 Nov;159(3):1117-34. doi: 10.1093/genetics/159.3.1117. Genetics. 2001. PMID: 11729157 Free PMC article.
-
RNA localization and translational regulation during axis specification in the Drosophila oocyte.Int Rev Cytol. 2001;203:541-66. doi: 10.1016/s0074-7696(01)03016-9. Int Rev Cytol. 2001. PMID: 11131526 Review.
-
Dorsoventral patterning in Drosophila oogenesis.Curr Opin Genet Dev. 1994 Aug;4(4):502-7. doi: 10.1016/0959-437x(94)90064-a. Curr Opin Genet Dev. 1994. PMID: 7950316 Review.
Cited by
-
The molecular tug of war between immunity and fertility: Emergence of conserved signaling pathways and regulatory mechanisms.Bioessays. 2020 Dec;42(12):e2000103. doi: 10.1002/bies.202000103. Epub 2020 Nov 9. Bioessays. 2020. PMID: 33169418 Free PMC article. Review.
-
Hydroxyurea exposure triggers tissue-specific activation of p38 mitogen-activated protein kinase signaling and the DNA damage response in organogenesis-stage mouse embryos.Toxicol Sci. 2013 Jun;133(2):298-308. doi: 10.1093/toxsci/kft069. Epub 2013 Mar 14. Toxicol Sci. 2013. PMID: 23492809 Free PMC article.
-
Expression of the Drosophila melanogaster GADD45 homolog (CG11086) affects egg asymmetric development that is mediated by the c-Jun N-terminal kinase pathway.Genetics. 2007 Nov;177(3):1691-702. doi: 10.1534/genetics.107.079517. Genetics. 2007. PMID: 18039880 Free PMC article.
-
A muscle-specific p38 MAPK/Mef2/MnSOD pathway regulates stress, motor function, and life span in Drosophila.Dev Cell. 2011 Oct 18;21(4):783-95. doi: 10.1016/j.devcel.2011.09.002. Dev Cell. 2011. PMID: 22014527 Free PMC article.
-
Anthrax lethal factor and edema factor act on conserved targets in Drosophila.Proc Natl Acad Sci U S A. 2006 Feb 28;103(9):3244-9. doi: 10.1073/pnas.0510748103. Epub 2006 Feb 2. Proc Natl Acad Sci U S A. 2006. PMID: 16455799 Free PMC article.
References
-
- Anderson KV. Pinning down positional information: Dorsal-ventral polarity in the Drosophila embryo. Cell. 1998;95:439–442. - PubMed
-
- Cano E, Mahadevan LC. Parallel signal processing among mammalian MAPKs. Trends Biochem Sci. 1995;20:117–122. - PubMed
-
- Christerson LB, McKearin DM. orb is required for anteroposterior and dorsoventral patterning during Drosophila oogenesis. Genes & Dev. 1994;8:614–628. - PubMed
-
- Clark I, Giniger E, Ruohola-Baker H, Jan LY, Jan YN. Transient posterior localization of a kinesin fusion protein reflects anteroposterior polarity of the Drosophila oocyte. Curr Biol. 1994;4:289–300. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous