

The versatility of paramyxovirus RNA polymerase stuttering
- PMID: 10364305
- PMCID: PMC112614
- DOI: 10.1128/JVI.73.7.5568-5576.1999
The versatility of paramyxovirus RNA polymerase stuttering
Abstract
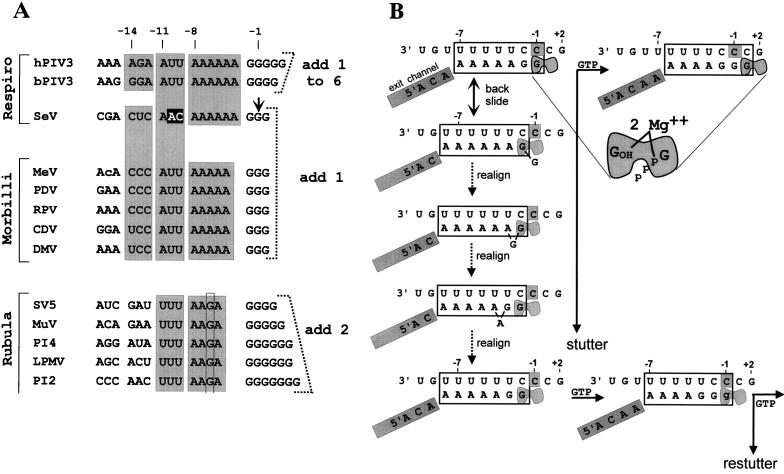
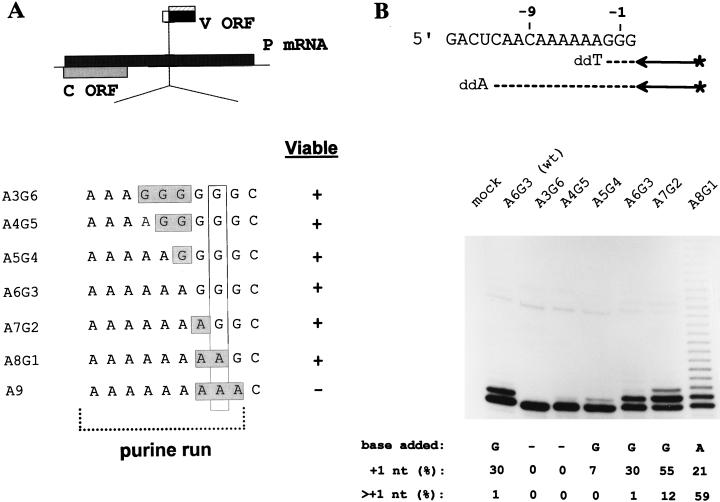
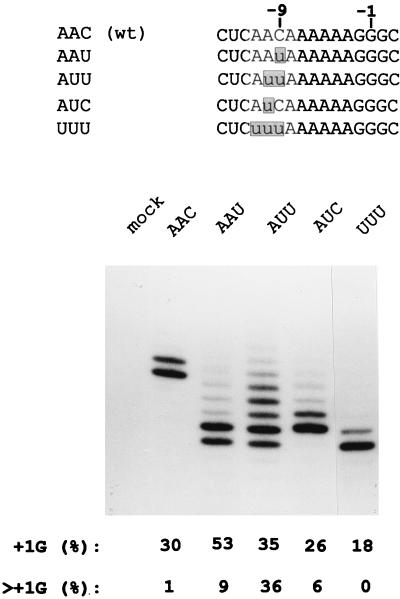
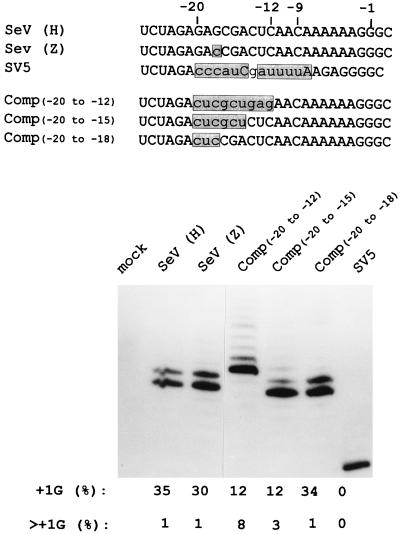
Paramyxoviruses cotranscriptionally edit their P gene mRNAs by expanding the number of Gs of a conserved AnGn run. Different viruses insert different distributions of guanylates, e.g., Sendai virus inserts a single G, whereas parainfluenza virus type 3 inserts one to six Gs. The sequences conserved at the editing site, as well as the experimental evidence, suggest that the insertions occur by a stuttering process, i.e., by pseudotemplated transcription. The number of times the polymerase "stutters" at the editing site before continuing strictly templated elongation is directed by a cis-acting sequence found upstream of the insertions. We have examined the stuttering process during natural virus infections by constructing recombinant Sendai viruses with mutations in their cis-acting sequences. We found that the template stutter site is precisely determined (C1052) and that a relatively short region (approximately 6 nucleotides) just upstream of the AnGn run can modulate the overall frequency of mRNA editing as well as the distribution of the nucleotide insertions. The positions more proximal to the 5' AnGn run are the most important in this respect. We also provide evidence that the stability of the mRNA/template hybrid plays a determining role in the overall frequency and range of mRNA editing. When the template U run is extended all the way to the stutter site, adenylates rather than guanylates are added at the editing site and their distribution begins to resemble the polyadenylation associated with mRNA 3' end formation by the viral polymerase. Our data suggest how paramyxovirus mRNA editing and polyadenylation are related mechanistically and how editing sites may have evolved from poly(A)-termination sites or vice versa.
Figures
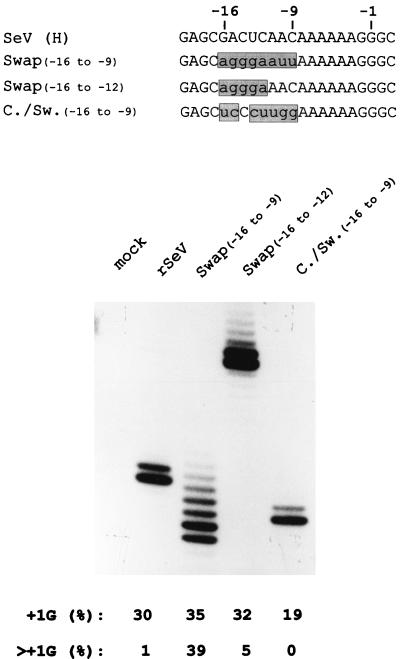
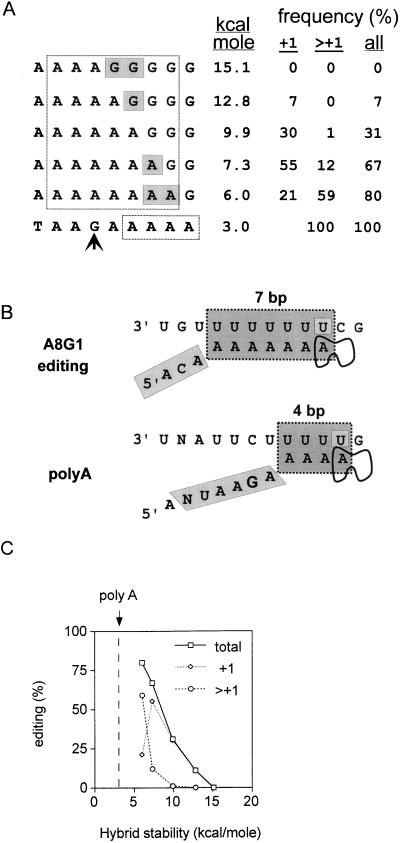
References
-
- Benne R. RNA editing: an overview. In: Benne R, editor. RNA editing. Chichester, England: Ellis Horwood; 1993. pp. 13–24.
-
- Benne R, Van den Burg J, Brakenhoff J P, Sloof P, Van Boom J H, Tromp M C, Major M C. Transcript of the frameshifted coxII gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA. Cell. 1986;46:819–826. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
