

Characterization of phosphorylation sites on the glutamate receptor 4 subunit of the AMPA receptors
- PMID: 10366608
- PMCID: PMC6782640
- DOI: 10.1523/JNEUROSCI.19-12-04748.1999
Characterization of phosphorylation sites on the glutamate receptor 4 subunit of the AMPA receptors
Abstract
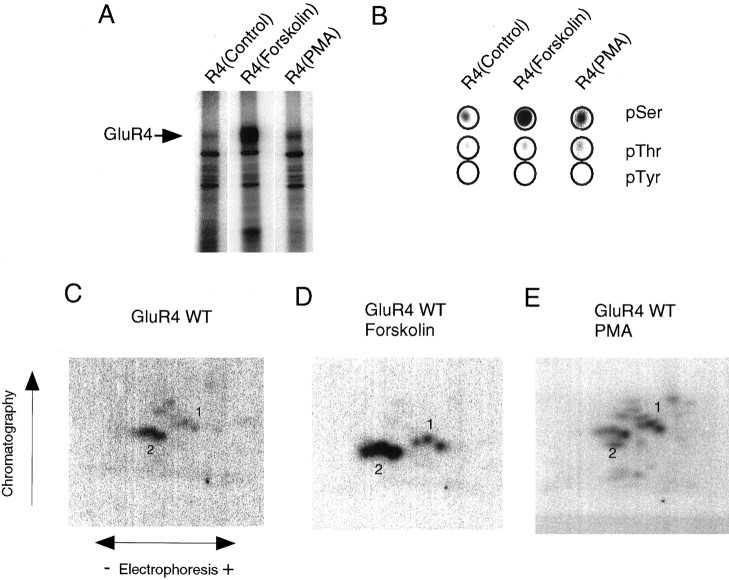
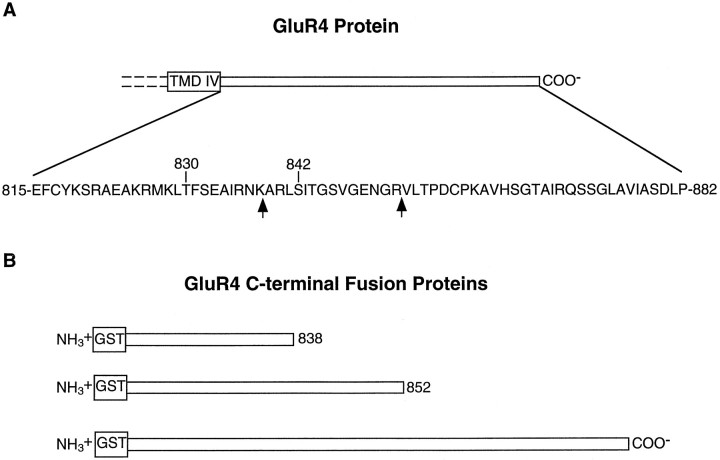
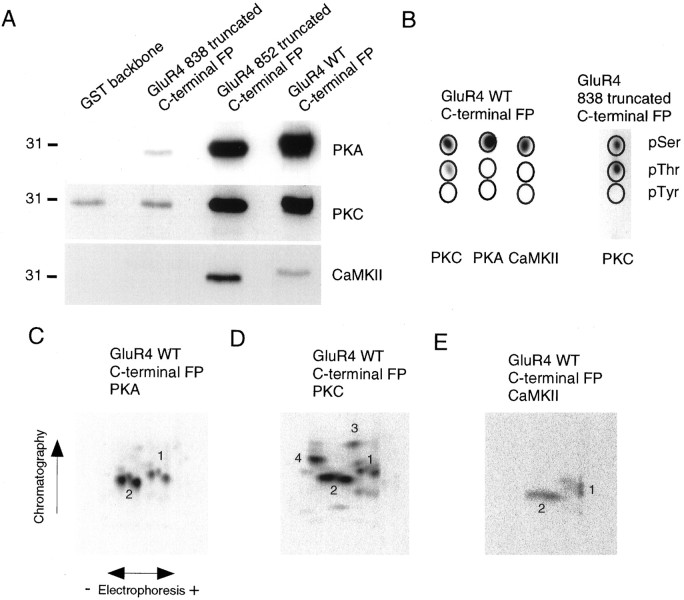
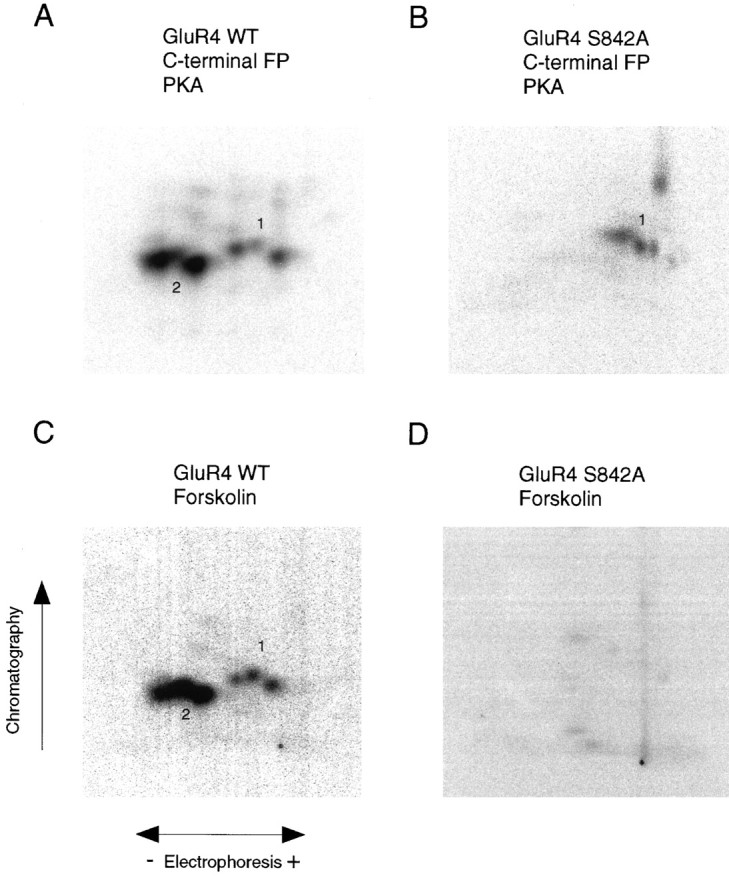
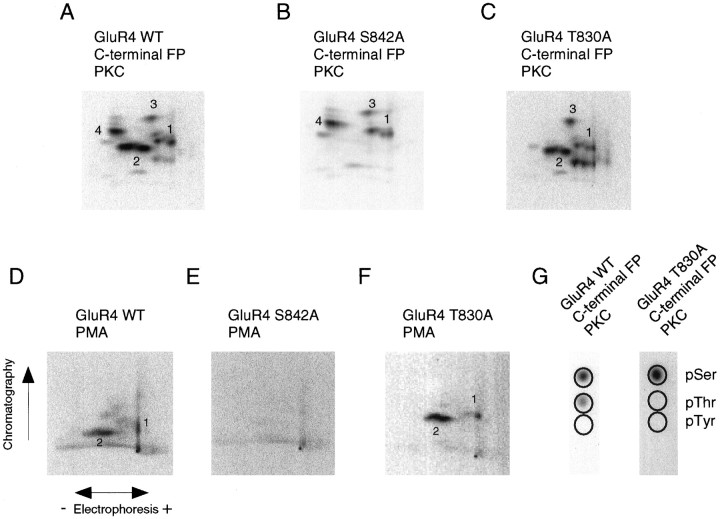
Recent studies have suggested that protein phosphorylation of glutamate receptors may play an important role in synaptic transmission. Specifically, the phosphorylation of AMPA receptors has been implicated in cellular models of synaptic plasticity. The phosphorylation of the glutamate receptor 1 (GluR1) subunit of AMPA receptors by protein kinase A (PKA), protein kinase C (PKC), and Ca2+/calmodulin-dependent protein kinase II (CaMKII) has been characterized extensively. Phosphorylation of this subunit occurs exclusively on the intracellular C-terminal domain. However, the GluR1 subunit C terminus shows low homology to the other AMPA receptor subunits. In this paper we characterized the phosphorylation of AMPA receptor subunit GluR4, using site-specific mutagenesis and biochemical techniques. We found that GluR4 is phosphorylated on serine 842 within the C-terminal domain in vitro and in vivo. Serine 842 is phosphorylated by PKA, PKC, and CaMKII in vitro and is phosphorylated in transfected cells by PKA. Two-dimensional phosphopeptide analysis indicates that serine 842 is the major phosphorylation site on GluR4. In addition, we identified threonine 830 as a potential PKC phosphorylation site. These results suggest that GluR4, which is the most rapidly desensitizing AMPA receptor subunit, may be modulated by phosphorylation.
Figures
References
-
- Barria A, Derkach V, Soderling T. Identification of the Ca2+/calmodulin-dependent protein kinase II regulatory phosphorylation site in the α-amino-3-hydroxy-5-methyl-4-isoxazole-propionate-type glutamate receptor. J Biol Chem. 1997a;272:32727–32730. - PubMed
-
- Barria A, Muller D, Derkach V, Griffith LC, Soderling TR. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science. 1997b;276:2042–2045. - PubMed
-
- Bennett JA, Dingledine R. Topology profile for a glutamate receptor: three transmembrane domains and a channel-lining reentrant membrane loop. Neuron. 1995;14:373–384. - PubMed
-
- Bliss TVP, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous